|
|
Ферменты биосинтеза ДНК. Прокариоты.Хеликаза – раскручивает двойную спираль ДНК в репликационной вилке. РНК-полимераза (праймаза) катализирует синтез олигорибонуклеотида (от 10 до 60 нуклеотидов), т. е. праймера, с которого начинается синтез ДНК. Праймасома - комплекс, включающий около 20 полипептидов. Участвует в формировании специфической вторичной структуры ДНК, подходящей для узнавания праймазой. ДНК-полимераза I катализирует отщепление праймера, удаление ошибочно присоединенных нуклеотидных остатков и заполнение образующихся пробелов (ДНК-полимеразная активность). ДНК-полимераза II достраиваете поврежденные участки в молекуле ДНК, т. е. осуществляет репарациюДНК. ДНК-полимераза III катализирует синтез ведущей (лидирующей) и отстающей цепей ДНК при репликации. ДНК-лигаза соединяет две цепи ДНК или замыкает два конца одной цепи в процессе репликации либо репарации. Топоизомеразы создают или уничтожают суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК. ДНК-полимеразы эукариот – α, β, γ, d, ε. В репликации ДНК участвуют два главных типа полимераз - α и d. ДНК-полимераза γ реплицирует митохондриальную ДНК. ДНК-полимераза ε в ряде случаев заменяет ДНК-полимеразу d. ДНК-лигаза устраняет разрывы в одной из цепей ДНК, замыкает линейную молекулы ДНК в кольцевую структуру. Белковые факторы, необходимые для биосинтеза ДНК. ДНК-связывающий белок (SSB-белок).Ослабляет взаимодействие цепей в молекуле ДНК. Активирует ДНК-полимеразы II и III. ДНК-раскручивающий белокобладает нуклеазной активностью. Разрывает связь одной из цепей ДНК, что обеспечивает раскручивание ее молекулы. ДНК-закручивающий белок вызывает суперспирализацию ДНК. У эукариот открыты факторы репликации – RFA (аналог SSB-белка) и RFC – фактор, стабилизирующий репликационный комплекс. Всего в репликации ДНК участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему,называемую реплисомой. Этапы биосинтеза ДНК: Инициация. К одноцепочечному фрагменту ДНК в момент распаривания биспиральной структуры присоединяются ДНК-связывающий белок, ДНК-раскручивающий белок, ДНК-полимеразный комплекс, праймаза и праймосома.Формируется репликативная вилка(рис. 9). На материнской цепи ДНК при участии праймазысоздается затравочный олигонуклеотид – праймер. Элонгация.При посредстве ДНК-полимеразы III на материнской цепи ДНК синтезируется дочерняя цепь. Процесс полимеризации идет по принципу комплементарности и только в направлении 5' ® 3'.Обе цепи реплицируются одновременно. Их синтез идет в противоположных направлениях.
Синтез ведущей цепи ДНК осуществляется непрерывно. Отстающая цепь образуется в направлении, обратном движению репликативной вилки. Синтез происходит фрагментарно. Эти фрагменты получили название фрагментов Оказаки (в честь японского биохимика, впервые предложившего схему биосинтеза ДНК, в которой были преодолены трудности, связанные с антипараллельностью цепей ДНК в ее биспиральной молекуле). Длина фрагментов Оказаки – 150-200 тысяч нуклеотидов у эукариот и 1000-2000 – у бактерий. Элонгация завершается отделением праймеров и замещением свободных мест комплементарными дезоксирибонуклеотидами под действием ДНК-полимеразы I. Отдельные фрагменты ДНК объединяются при помощи ДНК-лигаз. Если бы молекула ДНК реплицировалась одним репликативным комплексом, то потребовалось бы 10 дней. Фактически репликация генома человека in vivo продолжается 6-8 ч. Репликация одновременно начинается в нескольких участках ДНК, имеющих определенную нуклеотидную последовательность и называемых ориджинами. Ориджины расположены примерно через 100000 нм. Участок ДНК между соседними ориджинами называют репликоном.Каждый репликон реплицируется двумя репликативными комплексами, движущимися навстречу друг другу. Один репликон реплицируется за 2 ч. И столько же времени потребуется для репликации молекулы ДНК любой длины. Терминация.Существуют особые нуклеотидные последовательности, программирующие прекращение репликации ДНК. Точность репликацииДНК – одна ошибка на 1010 реакций. Допущенная ошибка может быть исправлена в ходе репарационных процессов. Синтез ДНК на матрице РНК В составе онковирусов (вирус саркомы Рауса и Раушера) открыт фермент обратная транскриптаза, или ревертаза (РНК-зависимая ДНК-полимераза), катализирующий биосинтез молекулы ДНК на матрице РНК. Ревертаза нуждается в праймере, роль которого может играть тРНК. На I этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК. Образуется так называмая гибридная молекула. II этап - разрушение исходной вирусной РНК. На IIIэтапе на матрице цепи ДНК комплементарно синтезируются новые цепи ДНК. Ревертаза нашла применение в молекулярной биологии для синтеза генов и фрагментов генов и в генетической инженерии для расшифровки первичной структуры РНК и белков.
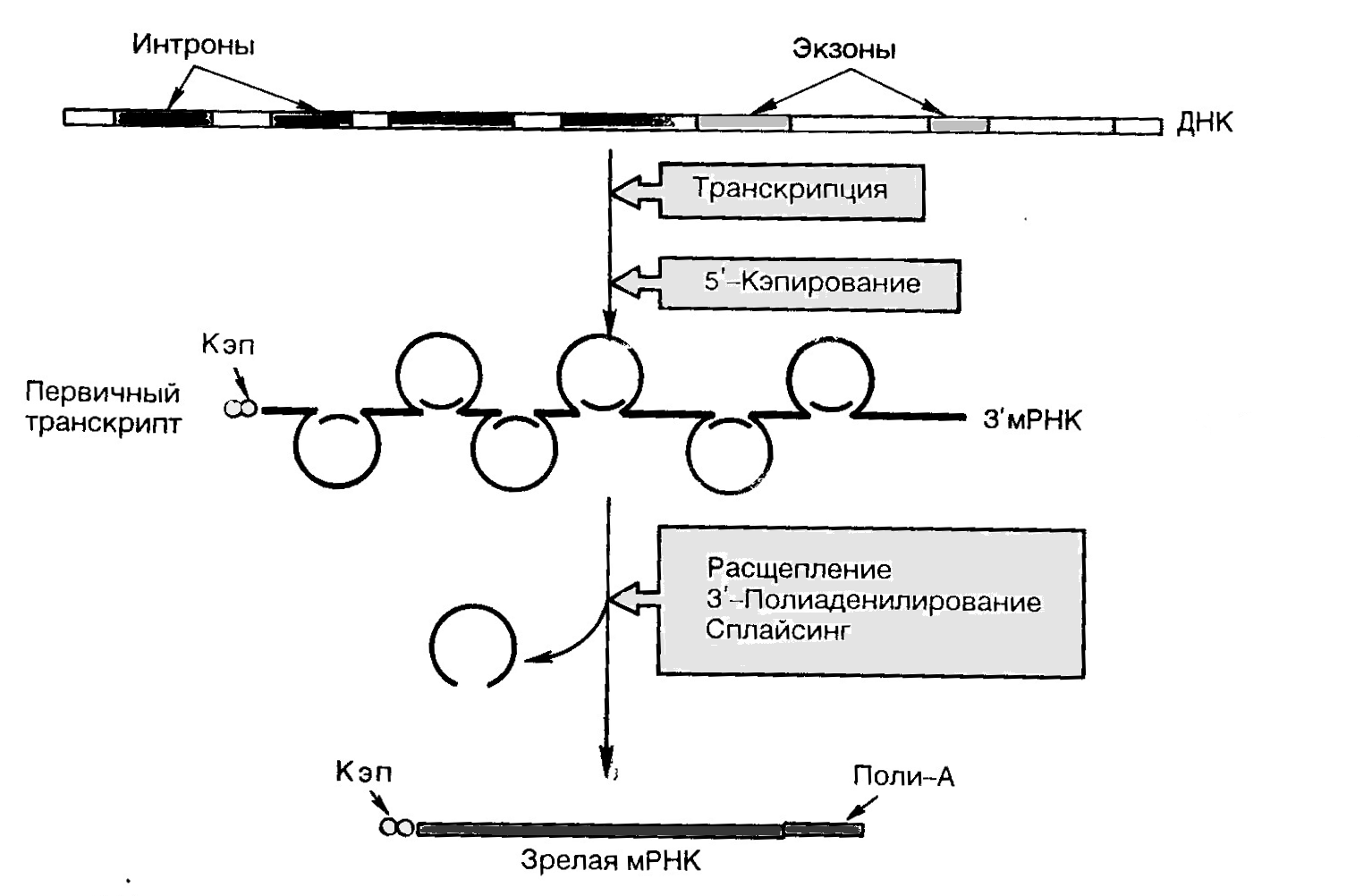
БИОСИНТЕЗ РНК Биосинтез РНК - это процесс транскрипции,т. е. переписывания информации с ДНК-матрицы.Биосинтез РНК осуществляется на ДНК при посредстве РНК-полимераз. У эукариототкрыты три разные РНК-полимеразы. РНК-полимераза I катализирует синтез рибосомных РНК, РНК-полимераза II - матричной РНК, РНК-полимераза III - транспортных РНК, а также ряда низкомолекулярных РНК со специфической функцией. Вместе с ферментом в единый транскрипционный комплекс объединены множество регуляторных белков (факторы транскрипции). Все виды РНК синтезируются на ядерной ДНК в качестве матрицы. Синтез мРНК.Одновременно на молекуле ДНК могут синтезироваться много молекул РНК. Ген эукариот наряду с кодирующими последовательностями (экзоны) содержит также некодирующие (интроны). РНК-полимераза катализирует транскрипцию как экзонов, так и интронов с образованием первичного транскрипта (РНК-предшественника). Наряду с информативными зонами они содержат неинформативные участки. В дальнейшем в ядре происходит процессинг(рис. 10),или созревание РНК. Основные этапы процессинга: 1. Кэпирование - химическая модификация 5'-концевой последовательности мРНК. «Колпачок», или «кэп», представляет собой 2. Полиаденилирование - химическая модификация 3'-концевой последовательности мРНК. Образование поли-А-последовательности длиной Кэп и поли-А-последовательность участвуют в инициации трансляции мРНК, а также защищают мРНК от гидролиза клеточными РНКазами. 3. Сплайсинг - удаление интронов из мРНК и сшивание образующихся экзонов. Данный процесс происходит при участии малых ядерных рибонуклеопротеинов. Такие РНК называют рибозимами. Это единственные из известных макромолекул, которые наделены как информационной, так и каталитической функцией. После завершения сплайсинга мРНК поступает в цитозоль.
Рис. 10. Процессинг мРНК
Первичные транскрипты тРНКпревращаются в зрелые формы путем сплайсинга и присоединения последовательности ЦЦА, в результате чего образуется акцепторный участок. Первичный транскрипт рРНКне содержит интронов, и при действии специфических РНКаз расщепляется с образованием более мелких молекул. Синтез РНК на матрице РНК Вирусная РНК индуцирует образование в клетках хозяина На I стадии РНК-репликаза на матрице РНК-вируса строит комплементарную цепь РНК. На II стадии она служит матрицей для синтеза РНК, однотипной исходной вирусной РНК. Обе стадии катализируются одним и тем же ферментом, хотя в каждой участвуют различные белковые факторы. Таким образом, поиск веществ, подавляющих активность
БИОСИНТЕЗ БЕЛКА Синтез белка тесно связан с понятием генетического кода. Генетический код - свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. Свойства генетического кода 1. Триплетность - единицей кода является сочетание трёх нуклеотидов (триплет, или кодон), всего 64 кодона, т.е. каждая аминокислота кодируется тремя нуклеотидами в мРНК (таблица 3). 2. Непрерывность - между триплетами нет знаков препинания, то есть информация считывается непрерывно. 3. Неперекрываемость - один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов. 4. Однозначность (специфичность) - определённый кодон соответствует только одной аминокислоте (АУУ - изолейцин, УУУ - фенилаланин). 5. Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов (изолейцин - АУУ, АУЦ, АУА). 6. Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусов до человека.
Таблица 3 Генетический код (и-РНК)
|
|