|
|
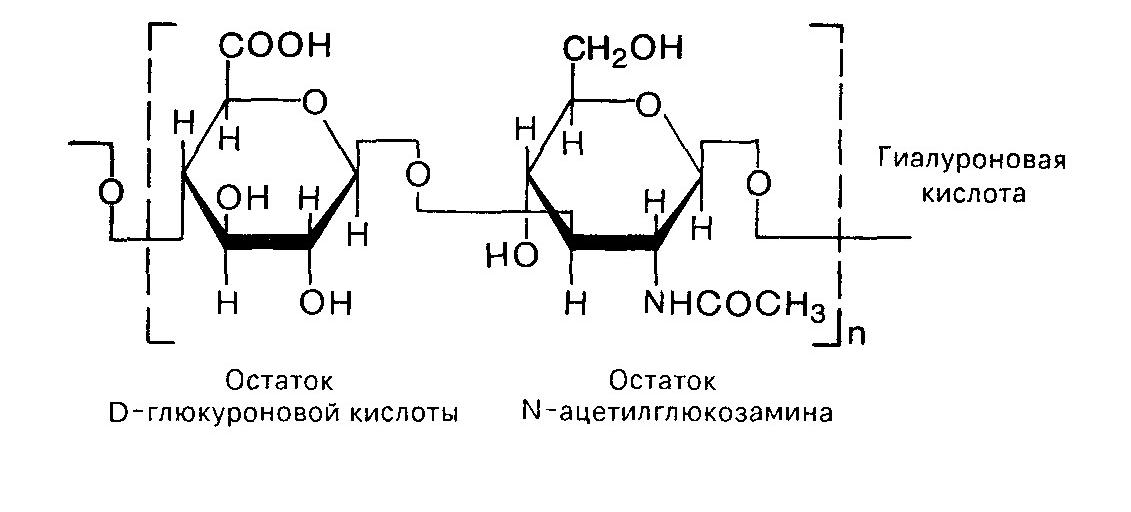
Неколлагеновые структурные гликопротеины.Фибронектин- димер. Его субъединицы соединены антипараллельно двумя дисульфидными связями в области С-концов. Пептидная цепь имеет несколько глобулярных доменов. Молекулы имеют вытянутую форму и имеют центры связывания с другими молекулами фибронектина, с коллагеном, гликозамингликанами, интегринами. Ламинин - гетеротример. Молекула ламинина имеет крестообразную форму, с тремя короткими ветвями, каждая из которых содержит глобулярные домены, и длинной ветвью с С-концевым глобулярным доменом. Ламинин в базальных мембранах находится в комплексе с другим белком - нидогеном. Нидоген имеет центры связывания коллагена. Он обеспечивает объединение основных компонентов базальной мембраны. В присутствии Са2+ ламинин путем самосборки образует сетевидные структуры за счет взаимодействия глобулярных доменов коротких ветвей. 4. Протеогликаны- высокомолекулярные углеводно-белковые соединения. Составляют до 30% от сухой массы соединительной ткани. Полисахаридная группа протеогликанов получила название мукополисахаридов (эти вещества обнаруживали преимущественно в слизистых субстратах). В дальнейшем их стали называть гликозаминогликанами.Это линейные неразветвленные полимеры, построенные из повторяющихся дисахаридных единиц. В их состав входят остатки глюкозамина, либо галактозамина, а также D-глюкуроновой или L-идуроновой кислот. Гиалуроновая кислотавпервые была обнаружена в стекловидном теле. Она построена из остатков глюкуроновой кислоты и
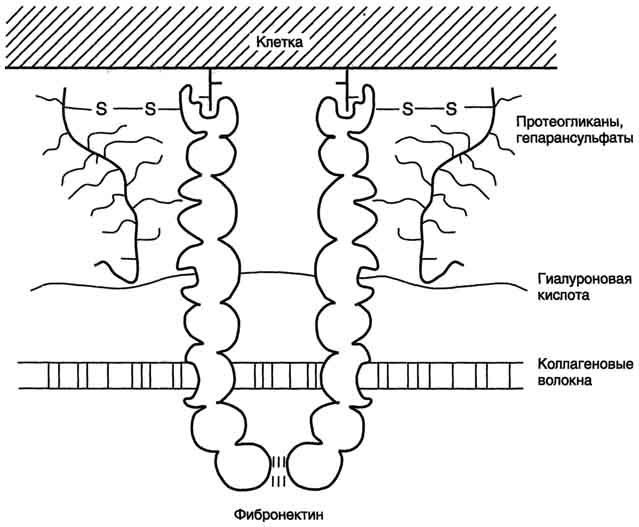
Хондроитинсульфаты содержат глюкуроновую кислоту и сульфированный N-ацетилгалактозамин. В сердечных клапанах содержится хондроитин-6-сульфат, в хряще - хондроитин-4-сульфат. Дерматансульфатсодержится в дерме (коже). Участвует в стабилизации коллагеновых пучков. В состав дерматансульфата входит L-идуроновая кислота. Кератансульфатвпервые был выделен из роговой оболочки глаза быка. Содержит сульфированный N-ацетиллактозамин. Кератансульфаты из разных тканей различаются по степени сульфатированности и типу связи с белком. Гепаринсинтезируется тучными клетками. Включает остатки Биосинтез гликозаминогликанов.Синтез глюкозамина и глюкуроновой кислоты происходит из D-глюкозы. Далее происходит эпимеризация глюкозамина в галактозамин, а глюкуроновой кислоты - в идуроновую кислоту. Предшественники гиалуроновой кислоты - уридиндифосфонуклеотидные производные N-ацетилглюкозамина и глюкуроновой кислоты. Белковая часть протеогликанов синтезируется на полирибосомах ЭПР. В полости ЭПР начинается синтез углеводной части. Здесь же происходит ее сульфирование. Одновременно с синтезом полисахаридных цепей молекула протеогликана продвигается по направлению к пластинчатому комплексу, где включается в секреторные гранулы и экзоцитируется. У разных протеогликанов белковые компоненты различны; они не имеют ничего общего с коллагеном и эластином. С одной пептидной цепью через гидроксильные группы серина связано большое число цепей гликозамингликана, так что молекула напоминает ершик. В межклеточном веществе протеогликаны содержатся главным образом в составе комплексов с гиалуроновой кислотой. Вследствие отталкивания отрицательно заряженных сульфатированных цепей гликозаминогликанов, а также их гидратации, молекулы в растворе по форме напоминают ершик. При увеличении давления (например, на хрящи суставных поверхностей) происходит «выдавливание» воды, и объем ткани уменьшается. Таким образом, протеогликаны выполняют функцию рессор в суставах. Интеграция межклеточного матрикса и клетокобеспечивается молекулами адгезии (рис. 31). Интегрины - трансмембранные белки, представляют собой димеры. Обеспечивают трансмембранную передачу сигнала. Обе цепи интегрина имеют большие внеклеточные домены и короткие внутриклеточные. Когда в матриксе в области внеклеточного домена интегрина происходят изменения, сигнал передается на внутриклеточный домен и регуляторные цитоплазматические белки, связанные с ним. Сигнализация интегринами снаружи внутрь приводит к изменению синтеза и секреции компонентов межклеточного матрикса или синтеза и секреции ферментов, разрушающих компоненты матрикса. Сходным образом происходит передача сигнала изнутри наружу.
Рис. 31. Интеграция компонентов межклеточного матрикса 10.2. ОСОБЕННОСТИ МЕТАБОЛИЗМА |
|