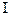|
|
Появление сходства корковых функциональных систем при мысленном и реальном выполнении бега у спринтера 1 разряда(по данным корреляционного анализа ЭЭГ)
Примечание: 1—8 — номера корковых зон, А — плеяды взаимосвязанной (синхронной и синфазной) активности различных корковых зон с коэффициентами корреляции 0.7-1.0, В — независимые корковые зоны. В процессах программирования используются имеющиеся у человека представления о «схеме тела», без которых невозможна правильная адресация моторных команд к скелетным мышцам в разных частях тела, и о «схеме пространства», обеспечивающие пространственную организацию движений. Нейроны, связанные с этими функциями, находятся в нижнетеменной ассоциативной области задних отделов коры больших полушарий. Организация движений во времени, оценка ситуации, построение последовательности двигательных актов, их сознательная целенаправленность осуществляются передне-лобной ассоциативной корой. Только в ней имеются специальные нейроны кратковременной памяти, которые удержи ва-ют созданную программу от момента прихода в кору внешнего пускового сигнала (или от момента самоприказа) до момента осуществления моторной команды. Соответственно этому во время реальной работы можно видеть особую специфику мозговой активности, отражающую характерные черты двигательных программ (рис. 32).Так, у бегунов и конькобежцев как при воображаемом,таки при реальном выполнении бега по
Рис. 32. Перестройка и специфика корковых функциональных систем на различных этапах выработки двигательных навыков а — мастера спорта, б — спортсмены-разрядники. На схемах — вид головы сверху. 1— иррадиация (б) и концентрация (а) медленных потенциалов в темпе движения («меченых ритмов») у бегунов во время бега; 2—5 — высокие межцентральные взаимосвязи потенциалов: 2— бегуны, бег; 3 — фехтовальщики, уколы с выпадом; 4 — биатлонисты, стрельба; 5 — тяжелоатлеты, толчок штанги.
дорожке или на коньках, устанавливается сходство (пространственная синхронизация) потенциалов передне-лобной (программирующей) области с моторными центрами ног, а у гимнастов при представлении и выполнении стойки на кистях — с моторными центрами рук. При стрельбе, бросках мяча в баскетбольное кольцо возникает сходство активности зрительных, нижнетеменных зон (ответственных за пространственную ориентацию движений) и моторных зон коры, что обеспечивает точность глазо-двигательных реакций. В процессе фехтования к этим зонам подключаются передне-лобные области, связанные с вероятностной оценкой текущей и будущей ситуации. В создании моторных программ принимают участие многие нейроны коры, мозжечка, таламуса, подкорковых ядер и ствола мозга. Обширное вовлечение множества мозговых элементов необходимо для поиска наиболее нужных из них. Этот процесс обеспечивается широкой иррадиацией возбуждения по различным зонам мозга и сопровождается обобщенным характером периферических реакций — их генерализацией. В силу этого первая стадия начинающихся попыток выполнить задуманное движение называется стадией генерализации. Она характеризуется напряжением большого числа активированных скелетных мышц, их продолжительным сокращением, одновременным вовлечением в движения мышц-антагонистов, отсутствием интервалов в ЭМГ во время расслабления мышц (рис. 33). Все это нарушает коорди нацию движений, делает их закреп ощенн ыми, при вод ит к значительным энерготратам и, соответственно, излишне выраженным вегетативным реакциям. На этой стадии наблюдаются особенное учащение дыхания и сердцебиения, подъем артериального давления, резкие изменение состава крови, заметное повышение температуры тела и потоотделения. Однако нет достаточной согласован ности этих сдвигов между собой и их соответствия мощности и характеру работы. Массированный поток афферентных импульсов от проприоре-цепторов многих мышц затрудняет отделение основных рабочих мышечных групп от посторонних. Анализ «темного» мышечного чувства еще более осложняется обильным притоком интероцептивных сигналов — в первую очередь, от рецепторов дыхательной и сердечно-сосудистой систем. Требуются многократные повторения разучиваемого упражнения для постепенного совершенствования моторной программы и приближения ее к заданному эталону. На второй стадии формирования двигательного навыка происходит концентрация возбуждения в необходимых для его осуществления корковых зонах. В посторонних же зонах коры активность подавляется одним из видов условного внутреннего торможения — дифференцировочным торможением. В коре и подкорковых структурах создается мозаика из возбужденных и заторможенных нейронных объединений, что обеспечивает координированное выполнение двигательного акта. Включаются лишь необходимые мышечные группы и только в нужные моменты движения, что можно видеть на записях ЭМГ. В результате рабочие энерготраты снижаются. Навык на этой стадии уже сформирован, но он еще очень непрочен и нарушается при любых новых раздражениях (выступление на незнакомом поле, появление сильного соперника и т. д.). Эти воздействия разрушают неокрепшую еще рабочую доминанту,
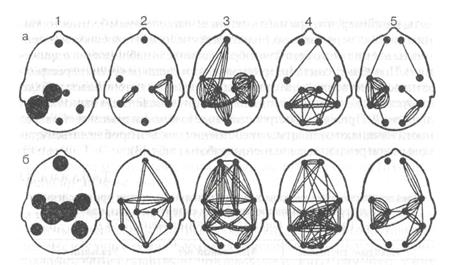
Рис. 33. Характеристика деятельности мышц при формировании двигательного навыка Электромиограмма трехглавой мышцы плеча при неосвоенных (А) и освоенных (Б) циклических движениях едва установившиеся межцентральные взаимосвязи в мозгу вновь приводят к иррадиации возбуждения и потере координации. На третьей стадии в результате многократного повторения навыка в разнообразных условиях помехоустойчивость рабочей доминанты повышается. Появляется стабильность и надежность навыка, снижается сознательный контроль за его элементами, т. е. возникает автоматизация навыка. Прочность рабочей доминанты поддерживается четкой сонастройкой ее нейронов на общий ритм корковой активности. Такое явление было названо А. А. Ухтомским усвоением ритма. При циклической работе ритм корковой активности соответствует темпу выполняемого движения: в ЭЭГпоявляются потенциалы, соответствующие этому темпу «меченые ритмы» ЭЭГ — рис. 34 (Сологуб Е.Б., 1965). Внешние раздражения на этой стадии лишь подкрепляют рабочую доминанту, не разрушая ее. Большая же часть посторонних афферентных потоков не пропускается в спинной и головной мозг: специальные команды из вышележащих центров вызывают пресинаптическое торможение импульсов от периферических рецепторов, препятствуя их доступу в спинной мозги вышележащие центры. Этим обеспечивается защита
Рис. 34. Медленные потенциалы в темпе движения — «меченые ритмы» ЭЭГ Радиотелеметрическая регистрация активности моторной области ноги левого полушария у спортсмена-спринтера при пробегании 50-метровых отрезков. Цифры справа — порядковые номера пробегов сформированных программ от случайных влияний и повышается надежность навыков. Процесс автоматизации не означает выключения коркового контроля за выполнением движения. В коре работающего человека отмечается появление связанных с движением потенциалов, специфические формы межцентральных взаимосвязей активности. Однако в этой системе центров по мере автоматизации снижается участие лобных ассоциативных отделов коры, что, по-видимому, и отражает снижение его осознаваемости. |
|