|
|
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ ТАМБОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ИМЕНИ Г.Р. ДЕРЖАВИНА
А.В. Гулин С.Е. Синютина А.Г. Шубина
БИОХИМИЯ (часть 2)
Допущено Редакционно-издательским советом ТГУ имени Г.Р. Державина в качестве учебного пособия для студентов, обучающихся по специальности 060101.65 – Лечебное дело 060103.65 – Педиатрия 060105.65 – Стоматология
Тамбов 2014
Рецензенты: Доктор медицинских наук, профессор Симонов С.Н.
СОдержАНИЕ
ОБМЕН ЛИПИДОВ Липиды– группа веществ, широко распространенных в живых организмах; нерастворимых в воде; растворимых в неполярных растворителях. Служат в организме источником энергии, выполняют термоизоляционную функцию, функцию защиты от механических воздействий, влияют на проницаемость биомембран, участвуют в передаче нервного импульса.
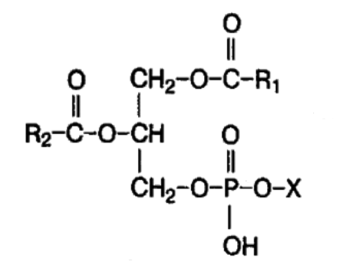
КЛАССИФИКАЦИЯ ЛИПИДОВ Жирные кислоты. Жирные кислоты организма человека содержат четное число атомов углерода. Существуют предельные и непредельные жирные кислоты. Непредельные представлены цис-изомерами. 2. Простые липиды -сложные эфиры жирных кислот с различными спиртами. 2.1. Триацилглицериныили жиры находятся в организме в форме протоплазматического жира, содержащегося в тканях в определенном количестве, и резервного жира, количество которого может значительно меняться. Жирные кислоты, входящие в их состав, содержат обычно от 12 до 24 атомов углерода, и, в основном, представлены пальмитиновой, стеариновой и олеиновой кислотами. 2.2. Воска - сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов с числом углеродных атомов от 16 до 22. 3. Сложные липиды -сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы. 3.1. Фосфолипидысодержат остаток фосфорной кислоты:
Примерами глицерофосфолипидовявляются: - фосфатидилхолины (лецитины) - содержат азотистое основание холин [НО—СН2—СН2—N+(CH3)3]; - фосфатидилэтаноламины -включают азотистое основание этаноламин НО—СН2—СН2—N+H3; - фосфатидилсерины -содержат остаток аминокислоты серина НО—СН2—СН(СОО-)—NH+3. Из всех липидов глицерофосфолипиды обладают наиболее выраженными полярными свойствами. Фосфатидилхолины и фосфатидилэтаноламины являются главными липидными компонентами мембран клеток. В составфосфатидилинозитоловвходит циклический спирт инозитол. Всфинголипидахспирт представлен сфингозином. Они находятся в мембранах клеток, особенно нервной ткани. Наиболее распространены сфингомиелины,содержащие остатки жирной кислоты, двухатомного ненасыщенного спирта сфингозина, азотистого основания (чаще холина) и фосфорной кислоты:
3.2. Гликолипиды (в основномгликосфинголипиды)широко представлены в нервной ткани. Содержат церамид, состоящий из спирта сфингозина и остатка жирной кислоты, и углеводы (чаще галактозу – галактозилцерамиды или глюкозу – глюкозилцерамиды).
Производные галактозилцерамидов – цереброзиды – могут выполнять рецепторные функции. 4. Стероидыотносятся к неомыляемой фракции липидов. Их представителями являются холестерин, стероидные гормоны, желчные кислоты, витамины группы D.
Каждая клетка содержит холестерин в составе мембран клеток. Он вместе с фосфолипидами и белками обеспечивает избирательную проницаемость клеточной мембраны и влияет на активность связанных с ней ферментов.
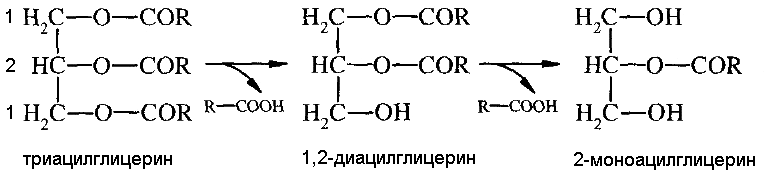
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ Суточная потребность в жирах составляет 50-100 г. Они обеспечивают до 50% потребности организма в энергии. У детей грудного возраста переваривание липидов начинается в ротовой полости. В секрете глоточных желез содержится лингвальная липаза, которая участвует в расщеплении эмульгированных жиров грудного молока. Оптимум рН для лингвальной липазы 4-5, что соответствует кислотности желудочного сока грудных детей. Основное переваривание жиров происходит в тонком кишечнике. В двенадцатиперстной кишке происходит нейтрализация попавшей в кишечник с пищей соляной кислоты желудочного сока бикарбонатами, содержащимися в панкреатическом и кишечном соках. Выделяющиеся при этом пузырьки углекислого газа способствуют перемешиванию пищевой кашицы. Происходит эмульгирование жира преимущественно солями желчных кислот, которые поступают в двенадцатиперстную кишку с желчью. В желчи содержатся холевая, дезоксихолевая и хенодезоксихолевая кислоты и небольшие количества других желчных кислот. Они образуют соли желчных кислот путем конъюгации с глицином или таурином. Поджелудочная железа секретирует неактивную пролипазу. Ее превращение в активную липазу происходит при участии желчных кислот и белка панкреатического сока - колипазы. Колипаза присоединяется к липазе и делает ее устойчивой к действию трипсина, а также способствует соединению липазы с мицеллами. Оптимум рН липазы 8-9, в присутствии желчи он смещается до 6. Липаза адсорбируется на поверхности мицелл и гидролизует сложноэфирную связь в триацилглицеринах. Основные продукты расщепления –
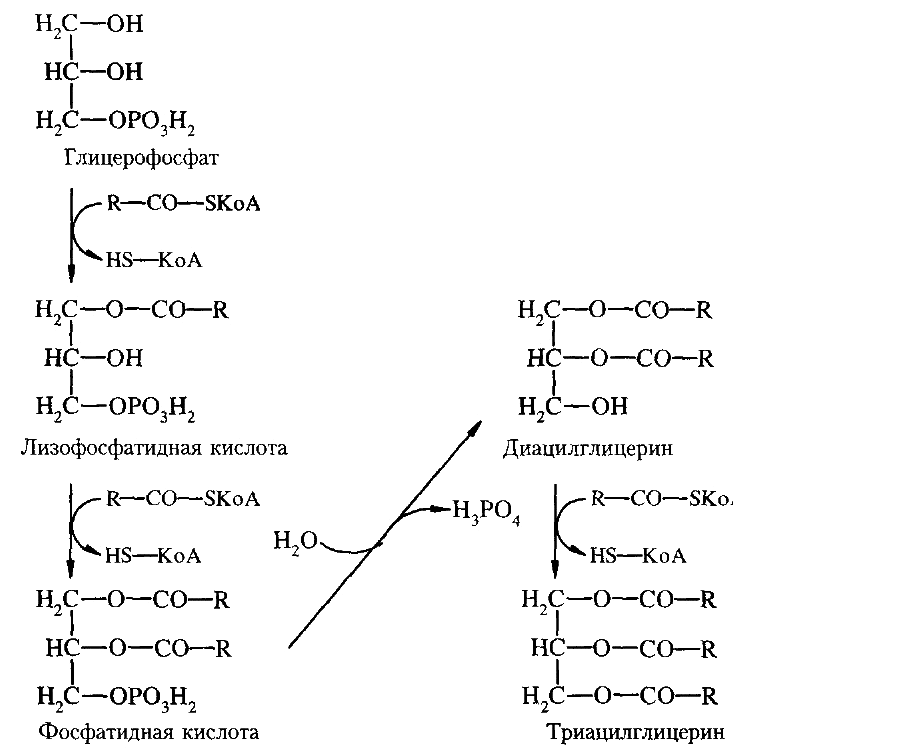
Панкреатический сок содержит фермент изомеразу, катализирующий превращение 2-моноацилглицерина в 1-моноацилглицерин, который затем гидролизуется липазой до жирной кислоты и глицерина. Жирные кислоты с короткой углеродной цепью и глицерин хорошо растворимы в воде. Они всасываются в кишечнике и поступают в кровь воротной вены, а затем в печень. Всасывание жирных кислот с длинной углеродной цепью и моноацилглицеринов происходит при участии желчи из мицелл. Жирные кислоты и моноацилглицерины всасываются, а соли желчных кислот остаются в просвете кишечника. Основная часть солей желчных кислот всасывается в кровь в подвздошной кишке, попадает в печень и затем выделяется с желчью. Этот процесс называется гепатоэнтеральной циркуляцией.За сутки он происходит 6-8 раз. В клетках эпителия кишечной стенки происходит ресинтез липидов. Он осуществляется двумя путями. Если в клетки поступили преимущественно жирные кислоты, синтез липидов идет глицерол-3-фосфатным путем, если жирные кислоты вместе с 2-моноацилглицеринами - 2-моноацилглиоловым путем. В 2-моноацилглицероловом пути из жирных кислот образуется их активная форма (ацил-КоА), затем происходит ацилирование 2-моноацилглицерин + R-CO-SKoA ® диацилглицерин + НS-KoA диацилглицерин + R-CO-SKoA ® триацилглицерин + НS-KoA. В глицерол-3-фосфатном пути на первом этапе глицерин фосфорилируется.Донором остатка фосфорной кислоты служит АТФ. Затем глицерол-3-фосфат взаимодействует с ацил-КоА. Далее процесс идет согласно схеме:
Липиды практически не растворимы в воде и в жидкостях организма. Поэтому необходимы специальные механизмы для их транспорта. Транспорт липидовосуществляется в составе особых сферических частиц – липопротеинов.Их поверхностная часть образована фосфолипидами, холестерином и белками (аполипопротеинами). Фосфолипиды гидрофильными концами ориентированы наружу, а гидрофобные концы находятся в липидной фазе внутри частиц. Внутренняя липидная фаза содержит в основном триацилглицерины и эфиры холестерина (рис. 1). Липопротеины различаются по составу и содержанию липидов и белков. Их плотность тем больше, чем больше содержание в них белков. Липопротеины разделяют центрифугированием и элеткрофорезом. - хиломикроны(ХМ) - диаметр – 300-500 нм; - липопротеины очень низкой плотности (ЛОНП) – 30-80 нм; - липопротеины низкой плотности (ЛНП) – 20-25 нм: - липопротеины высокой плотности (ЛВП) – 5-12 нм.
Липопротеины образуются в клетках слизистой оболочки кишечника (хиломикроны и ЛОНП), в плазме крови (ЛНП и ЛВП), в гепатоцитах (ЛОНП и ЛВП). Основные функции хиломикроны и ЛОНП – транспорта жиров по кровеносному руслу, а ЛНП и ЛВП – транспорт холестерина. Жиры, синтезирующиеся в клетках кишечника, включаются главным образом в ХМ, и небольшое количество – в ЛОНП, и попадают в лимфатическую систему и затем в общий кровоток. Через 1-2 ч после приема жирной пищи возникает алиментарная гиперлипемия. Это физиологическое явление, когда концентрация тригациллицеринов в крови повышается, и в ней появляются ХМ. Ее пик наблюдается через 4-6 часов, а через 10-12 ч содержание триацилглицеринов возвращается к норме. ХМ исчезают из крови. Триацилглицерины хиломикронов гидролизуются на поверхности эндотелия капилляров жировой ткани при участии фермента липопротеинлипазы. Образуются жирные кислоты и глицерин. Глицерин транспортируется в печень, где может быть использован для синтеза жиров. Основная масса жирных кислот проникает в ткани. В жировой ткани они депонируются в виде жиров, в сердечной мышце и работающих скелетных мышцах используются как источник энергии. Остаточные хиломикроны поглощаются клетками печени. |
|


 галактозилцерамид
галактозилцерамид холестерин
холестерин