|
|
Механизм естественного отбораВ процессе естественного отбора закрепляются мутации, увеличивающие приспособленность организмов. Естественный отбор часто называют «самоочевидным» механизмом, поскольку он следует из таких простых фактов, как: 1. Организмы производят потомков больше, чем может выжить; 2. В популяции этих организмов существует наследственная изменчивость; 3. Организмы, имеющие разные генетические черты, имеют различную выживаемость и способность размножаться. Такие условия создают конкуренцию между организмами в выживании и размножении и являются минимально необходимыми условиями для эволюции посредством естественного отбора. Таким образом, организмы с наследственными чертами, которые дают им конкурентное преимущество, имеют большую вероятность передать их своим потомкам, чем организмы с наследственными чертами, не имеющими подобного преимущества. Формы естественного отбора Существуют разные классификации форм отбора. Широко используется классификация, основанная на характере влияния форм отбора на изменчивость признака в популяции. Движущий отбор — форма естественного отбора, которая действует при направленном изменении условий внешней среды. Описали Дарвин и Уоллес. В этом случае особи с признаками, которые отклоняются в определённую сторону от среднего значения, получают преимущества. При этом иные вариации признака (его отклонения в противоположную сторону от среднего значения) подвергаются отрицательному отбору. В результате в популяции из поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию). Стабилизирующий отбор — форма естественного отбора, при которой его действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака. Понятие стабилизирующего отбора ввел в науку и проанализировал И. И. Шмальгаузен. Дизруптивный (разрывающий) отбор — форма естественного отбора, при котором условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дарвин описывал действие дизруптивного отбора, считая, что он лежит в основе дивергенции, хотя и не мог привести доказательств его существования в природе. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования. Половой отбор — это естественный отбор на успех в размножении. Выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Дарвин назвал это явление половым отбором. «Эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола». Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения, значительно выше, чем их недостатки для выживания. Распространены две гипотезы о механизмах полового отбора. · Согласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков. · Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности.
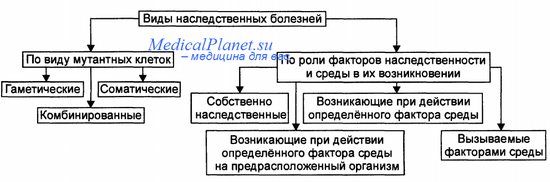
№7 Дизруптивный отбор Дизруптивный (разрывающий) отбор — форма естественного отбора, при котором условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дарвин описывал действие дизруптивного отбора, считая, что он лежит в основе дивергенции, хотя и не мог привести доказательств его существования в природе. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования. Одна из возможных в природе ситуаций, в которой вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам. Примером дизруптивного отбора является образование двух рас у погремка большого на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая. Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись только особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора. Билет№24 №1 диагностика дикроцелиоза Неоднократное обнаружение в дуоденальном содержимом и фекалиях яиц ланцетовидного сосальщика. №2 как связан КГ с ЭПС : В цис компартминт КГ поступают белки и липиды из цистерн ГЭПС , далее везикулы транспортируют их в промежуточный компартмент , затем в транс , из которого готовые продукты выходят в транс сеть АГ в котором происходит сортировка и специализация готовых молекул. №3 геномная мутация Геномные мутации - это мутации, которые приводят к добавлению либо утрате одной, нескольких или полного гаплоидного набора хромосом. Разные виды геномных мутаций называют гетероплоидией и полиплоидией. Полиплоидия — увеличение числа наборов хромосом, кратное гаплоидному (Зn, 4n, 5n и т.д.). Причины: двойное оплодотворение и отсутствие первого мейотического деления. У человека полиплоидия, а также большинство анеуплоидий приводят к формированию леталей. Анеуплоидия — изменение (уменьшение — моносомия, увеличение — трисомия) числа хромосом в диплоидном наборе, т.е. не кратное гаплоидному (2n+1, 2n-1 и т.д.). Механизмы возникновения: нерасхождение хромосом (хромосомы в анафазе отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и «анафазное отставание» (в анафазе одна из передвигаемых хромосом отстаёт от всех других). Трисомия — наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау). Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0). Подходы к классификации наследственных болезней приведены на рисунке. Основные критерии классификации наследственных болезней: вид мутантных клеток и вклад наследственных факторов и среды. В зависимости от вида первично поражённых клеток вьщелены следующие группы заболеваний: • болезни вследствие мутаций в половых клетках — «гаметические», т.е. собственно наследственные болезни (например, фенилкетонурия, гемофилии); эти болезни передаются по наследству; • болезни вследствие мутаций в соматических клетках — «соматические» (например, опухоли, некоторые болезни иммунной аутоагрессии). Эти заболевания не передаются по наследству; • болезни вследствие комбинации мутаций в половых и соматических клетках (например, семейная ретинобластома). Отдельно рассматривают летальные, сублетальные и гипогенитальные заболевания: • летальные болезни приводят к гибели во время внутриутробного развития (например, моносомии по аутосомам, гаплоидии, большинство полиплоидии); • сублетальные заболевания приводят к гибели индивида до периода полового созревания (например, наследственные иммунодефициты типа агам-маглобулинемии швейцарского типа, синдром Луи-Бар, некоторые гемофилии); • гипогенитальные заболевания сочетаются с бесплодием (например, синдромы Шерешевского—Тернера, Кляйнфелтера). В зависимости от роли наследственности и среды выделяют четыре группы заболеваний: • собственно наследственные болезни (развиваются вследствие мутаций); • экогенетические заболевания (развиваются при наличии мутации под влиянием специфического фактора среды); • болезни генетической предрасположенности (вклад генетических факторов очевиден); • болезни, вызываемые факторами внешней среды (строго говоря, эта группа болезней не относится к наследственным болезням, так как вклад генетических факторов в их возникновение, развитие и исход может быть крайне малым, сомнительным или неизвестным).
№4 к чему присоедин хромосомы (Центромером) №5 географическое распространение анкилостома Анкилостома (Ancylostoma duodenale) – возбудитель анкилостомоз. Географическое распространение: страны с тропическим климатом, преимущественно между 36 ° с. ш. и 30 ° ю. ш. В странах с умеренным климатом ячейки анкилостомоз встречаются в шахтах, где высокая влажность и постоянная температура благоприятные для развития личинок. №6 транскрипция у прокариот РНК-полимераза прокариот обеспечивает транскрипцию генов, несущих информацию о последовательностях молекул РНК всех трех классов: рибосомной РНК (рРНК), транспорт-ной РНК (тРНК) и информационной (или матричной) РНК (мРНК). №7 миграция как фактор эволюции Генетические различия между локальными популяциями возникают под действием разных факторов эволюции.Каждый вид живых организмов можно рассматривать как группу локальных популяций. Эти популяции часто отличаются друг от друга по генетическому составу. Каждая конкретная мутация является редким событием. Поэтому каждая популяция имеет свой, характерный только для нее, набор мутантных аллелей. Все популяции одного вида живут в сходных условиях. Однако для видов, широко распространенных, эти условия оказываются несколько различными в разных частях ареала. Эти различия касаются как абиотических факторов, таких как особенности климата, химического состава почвы и воды, так и биотических (набор и численность хищников и паразитов, видов конкурентов и др.). Естественный отбор приспосабливает каждую популяцию к локальным условиям среды. Так, например, в городских популяциях березовой пяденицы он приводит к повышению частоты темных форм, а в сельских – к ее понижению. В популяциях человека, населяющих малярийные районы, отбор поддерживает высокую частоту различных мутантных форм гемоглобина, а в других районах он эти формы элиминирует. Тем самым естественный отбор усиливает генетические различия между популяциями. Миграция нивелируют генетические различия между популяциями.Популяции одного вида, как правило, не изолированы друг от друга. Между ними постоянно происходит обмен генами. Животные из одной популяции переселяются в другую. Пыльца и семена растений переносятся ветром, течениями, животными из одной популяции в другую. Интенсивность обмена генами между популяциями зависит от расстояния между ними, радиуса индивидуальной активности у животных и распространения пыльцы и семян у растений и многих других причин. В результате обмена мигрантами между двумя соседними популяциями, отличающимися по генетическому составу, частоты аллелей в каждой из них меняются из поколения в поколение. Эти изменения тем более значительны, чем больше различия между популяциями в частотах аллелей и интенсивнее миграция. Поскольку мы определили эволюцию как изменение частот аллелей в популяциях, мы можем рассматривать миграцию как фактор эволюции. Эволюционным последствием миграции и обмена генами является нивелировка генетических различий между локальными популяциями. Таким образом, миграция как фактор эволюции, противодействует таким дифференцирующим популяции факторам – отбору, дрейфу генов и мутационному процессу. В этом смысле она тормозит эволюцию. Естественный отбор приспосабливает популяцию к локальным условиям существования, оставляя в ее генетическом составе наиболее адаптивные аллели. Миграция этому противодействует, постоянно поставляя в популяцию менее приспособленные к той среде, в которой живет эта популяция, варианты генов. Светлые сельские формы березовой пяденицы, мигрируя в города и вливаясь в состав локальных популяций, снижают их среднюю приспособленность. Благодаря миграциям поддерживается генетическое единство вида.Мутация, возникшая в любой из локальных популяций, имеет шанс распространиться на весь вид в целом. Мутации возникают случайно. Мутация, обеспечивающая устойчивость насекомых к инсектицидам, с равной вероятностью может возникнуть и в той популяции, которая обрабатывается инсектицидами, и в той, которая такому воздействию не повергается. Благодаря миграциям, новые мутации проникают в те популяции, где они оказываются полезными. Исследуя устойчивость насекомых к инсектицидам, ученые обнаружили, что генетические механизмы устойчивости оказались неодинаковыми в различных популяциях. В одних случаях устойчивость определялась доминантным геном, в других — рецессивным, отмечено не только аутосомное наследование, но и наследование, сцепленное с полом. Обнаружены случаи полигенного и цитоплазматического наследования. Соответственно и физиологические механизмы устойчивости к инсектицидам оказались различными. Среди них: накопление яда кутикулой; повышенное содержание липидов, способствующих растворению инсектицида; повышение устойчивости нервной системы к действию ядов; снижение двигательной активности и др. Разные популяции оказываются устойчивы к разным типам инсектицидов. Эти адаптации – в каждой популяции своя - подхватываются и усиливаются естественным отбором. Благодаря обмену мигрантами популяции могут обмениваться своими приспособлениями. В результате под влиянием миграции и естественного отбора формируется общая для вида многокомпонентная система устойчивости к целому спектру инсектицидов. Обмен генами между популяциями поставляет материал для естественного отбора, который работает над тем, чтобы свести частные локальные адаптации воедино и тем самым повысить приспособленность их носителей. Таким путем создается общий генофонд вида, в котором аллели отдельных генов подобраны по их совместимости друг с другом.
БИЛЕТ 28 №1 лаб. диагн. трихоцефалеза Ответ: обнаружение в фекалиях яиц власоглава. №2 где расположены аллели Аллели — различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных хромосом и определяющие альтернативные варианты развития одного и того же признака. В диплоидном организме может быть два одинаковых аллели одного гена, в этом случае организм называется гомозиготным, или два разных, что приводит к гетерозиготному организму. №3 в какой стадии происходит разрушение ядерной оболочки Ответ: (профаза) №4 что такое гетерохроматин? Гетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость. Гетерохроматин– транскрипционно неактивный и конденсированный хроматин интерфазного ядра. Гетерохроматин располагается преимущественно по периферии ядра и вокруг ядрышек, состовляет 10% от общего хроматина. Типичный пример гетерохроматина – тельце Барра. (по гистологии) №5 модификационная изменчивость Модификационная (фенотипическая) изменчивость — изменения в организме, связанные с изменением фенотипа вследствие влияния окружающей среды и носящие, в большинстве случаев, адаптивный характер. Генотип при этом не изменяется. Модификационная изменчивость - это эволюционно закрепленные реакции организма на изменения условий внешней среды при неизменном генотипе. №6 абиотические факторы Абиотические факторы среды - компоненты и явления неживой, неорганической природы, прямо или косвенно воздействующие на живые организмы: климатические, почвенные и гидрографические факторы. №7 разрушение мембраны (не знаю, правильно написал или нет!!!) В процессе старения организма в клетке происходят постепенные и необратимые изменения. Прежде всего, существенно нарушается мембранная строение клетки. Поражается как наружная клеточная мембрана, так и мембраны эндоплазматической структурных элементов клетки. Разрушение мембран лизосом приводит к выходу ферментов в цитоплазму, повышение активности гидролаз, что приводит разрушение структур ДНК и гибель клетки. Значительно изменяется проницаемость клеточных мембран для различных веществ, прежде нарушаются механизмы активного транспорта, облегченной и обменной диффузий. №8 распростр малярии. Малярия широко распространена во всем тропическом и субтропическом поясе, а также в умеренной зоне - везде, где имеются условия для развития комаров рода Anopheles, личинки которых живут в мелких пресноводных водоемах
Билет №29
№11 Комбинативная изменчивость — изменчивость, которая возникает вследствие рекомбинации генов во время слияния гамет. Основные причины: · независимое расхождение хромосом во время мейоза; · случайная встреча половых гамет, а вследствие этого и сочетания хромосом во время оплодотворения; · рекомбинация генов вследствие кроссинговера. Билет №30 №1Лаб диагностика аскаридоза Достоверное установление аскаридоза основано на обнаружении личинок аскарид в мокроте и специфических антител к акаридам в анализе крови. На рентгенограмме легких в этой стадии могут выявляться инфильтраты. В кишечной стадии заболевания основным методом является исследование кала на яйца аскарид. Дополнительную информацию дает рентгеновское исследование кишечника с контрастом и УЗИ брюшной полости №2Митохондрия Их называют «энергетическими станциями клетки». Основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии в синтезе молекул АТФ, который происходит за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны. |
|