|
|
Атавистические пороки развития
Различного рода нарушения эмбриогенеза могут привести к формированию у высокоорганизованных организмов и человека таких признаков, которые при нормальных условиях у них не встречаются, но присутствуют у более или менее отделенных предков. Такие признаки называют атавизмами. Если они снижают жизнеспособность и проявляются как морфологические аномалии, то их называют атавистическими или анцестральными (от франц. ancetre — предок) пороками развития. По механизмам формирования различают три варианта атавизмов. Наиболее часто встречаются атавизмы, связанные с недоразвитием органов на тех этапах морфогенеза, когда они рекапитулировали предковое состояние. К примерам аномалий такого рода относятся двух- и трехкамерное сердце, гипоплазия, или недоразвитие диафрагмы, срединная расщелина твердого нёба, или «волчья пасть», и др. Атавизмы другого рода — результат нарушения редукции — персистирование (сохранение) и дальнейшее развитие эмбриональных структур, также рекапитулирующих морфологию, характерную для предков. К ним относят персистирование боталлова протока и правой дуги аорты (см. § 14.4), наличие ребер, связанных с шейным отделом позвоночника, боковые свищи шеи (см. разд. 14.2.1, 14.3.4). Третий тип атавистических пороков развития возникает в связи с нарушением перемещения органов в онтогенезе, результатом чего является их расположение в тех частях тела, где при нормальных условиях они находятся у предковых форм. У человека широко известны тазовое расположение почек (см. разд. 14.5.1), крипторхизм (неопущение яичника), высокое стояние плечевого пояса и др. Во всех случаях атавизмов ведущими механизмами их возникновения являются не обратные мутации, приводящие к формированию предкового фенотипа, а, вероятно, мутации регуляторных генов, которые контролируют скорость морфогенеза и запуск процессов, направленных на редукцию органов. 57. Основные эволюционные преобразования начального отдела пищеварительной системы позвоночных.
58. ФИЛОГЕНЕЗ ВИСЦЕРИАЛЬНОГО ЧЕРЕПА У ПОЗВОНОЧНЫХ ЖИВОТНЫХ. Череп позвоночных состоит из двух основных отделов - осевого и висцерального. 1. Осевой - черепная коробка (мозговой череп - neurocranium) - продолжение осевого скелета, служит для защиты головного мозга и органов чувств. 2. Висцеральный - лицевой (splanchnocranium), образует опору для передней части пищеварительного тракта.
Первая дуга у большинства современных позвоночных приобретает функцию челюстного аппарата - ее называют челюстной, а вторую - тоже по функции - подъязычной или гиоидной. Остальные, начиная с третьей и до седьмой, называют жаберными, т.к. они служат опорой для жаберного аппарата. На ранних этапах развития висцеральный и осевой череп не связаны друг с другом, позднее эта связь возникает. Общие для всех зародышей позвоночных закладки семи висцеральных дуг в процессе эмбрионального развития претерпевают у представителей разных классов соответственно и различные специфические изменения.
I. Низшие рыбы (хрящевые)- Chondrichthyes 1-я, она же челюстная дуга, состоит из двух крупных хрящей, вытянутых в переднезаднем направлении: верхнего - небноквадратного - первичная верхняя челюсть, нижнего - меккелева - первичная нижняя челюсть; они сзади сращены между собой и выполняют функцию первичной челюсти. 2-я, она же подъязычная, или гиоидная дуга состоит из следующих компонентов: 1) из двух, расположенных, вверху гиомандибулярных хрящей, которые сверху соединены с черепной коробкой, снизу- с гиоидом, а впереди - с челюстной дугой - первичной верхней челюстью; 2) из двух гиоидов, расположенных ниже гиомандибулярных хрящей, которые с ними и соединены; кроме того, гиоиды соединены с первичной нижней челюстью; 3) из непарной копулы (небольшой хрящ, соединяющий оба гиоида между собой). Исходя из расположения гиомандибулярного хряща, понятно, что ему принадлежит роль подвеска, соединяющего челюстную дугу с черепом. Такой тип соединения называется гиостилией, а череп - гиостильным.Это свойственно низшим позвоночным - всем рыбам. Остальные висцеральные дуги с третьей по седьмую образуют опору для дыхательного аппарата. II.Высшие рыбы - (костные) Osteichthyes. Основное отличие касается только челюстной дуги: 1) верхний элемент челюстной дуги (верхняя челюсть) состоит вместо одного крупного небноквадратного хряща из пяти элементов - небного хряща, квадратной кости и трех крыловидных хрящей; 2)впереди от первичной верхней челюсти образуются две крупные накладные кости, снабженные крупными зубами, - эти кости и становится - вторичными верхними челюстями; 3)дистальный конец первичной нижней челюсти тоже покрывает большая зубная кость, которая выдается далеко вперед и образует вторичную нижнюю челюсть. Подъязычная дуга сохраняет прежнюю функцию, т.е. череп остается гипостильным.
59. Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху. Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи. Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения. Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).
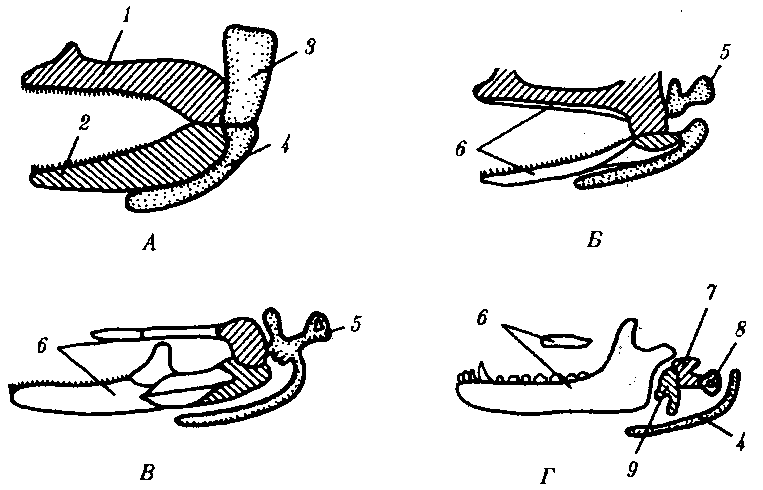
Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных. А—хрящевая рыба; Б—земноводное; В—пресмыкающееся; Г— млекопитающее: 1—нёбно-квадратный хрящ, 2—меккелев хрящ, 3—гиомандибулярный хрящ, 4—гиоид, 5—столбик, 6—накладные кости вторичных челюстей, 7—наковаленка, 8—стремечко, 9—молоточек; гомологичные образования обозначены соответствующей штриховкой Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
60. У Рептилий Костный. Сводчатое первичное нёбо. Вторичное нёбо образовано путем срастания нёбных отростков верхнечелюстных, межчелюстных костей и нёбных костей. Гиомандибулярный хрящ преобразовался в столбик, гиоид - в подъязычный аппарат, а жаберные дуги - в хрящи гортани. У Млекопитающих Костный. Верхняя челюсть образована сросшимися пред- и верхнечелюстными костями. Нижняя челюсть образована одной зубной костью, подвижно сочлененной с черепом. Верхний отдел челюстной дуги преобразуется в наковальню, а нижний - в молоточек, верхний отдел подъязычной дуги - в стремечко, а нижний - в передние рожки подъязычной кости. Первая жаберная дуга и ее копула преобразовались в тело подъязычной кости, задние рожки, а вторая и третья - в щитовидный хрящ.
Онто-филогенетические пороки висцерального черепа у человека. Щелевые дефекты губ, челюстей и нёба; вычленение и выдвижение вперёд межчелюстного (резцового) отдела верхней челюсти; прорезывание сверхкомплектных зубов и особенно резцов и премоляров; скученность зубов; сохранение трем обезьяних промежутков; выступающие сильноразвитые клыки; недифференцированные зубы, нарушающие правильный прикус. Получило объяснение частое сочетание пороков зубочелюстной систем с пороками органов слуха, что получило в клинике название синдромов 1-ой и 2-ой жаберных дуг.
61. Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную. У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части. Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю. У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными. ***Протостилия---подвижное соединение верх, челюсти с мозговым черепом без участия подъязычной дуги; наиб, примитивная форма подвески челюстей к черепу у позвоночных. П. существовала у древнейших рыб из группы акантод*** У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации. Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху. Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи. Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения. Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).
Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных. А—хрящевая рыба; Б—земноводное; В—пресмыкающееся; Г— млекопитающее: 1—нёбно-квадратный хрящ, 2—меккелев хрящ, 3—гиомандибулярный хрящ, 4—гиоид, 5—столбик, 6—накладные кости вторичных челюстей, 7—наковаленка, 8—стремечко, 9—молоточек; гомологичные образования обозначены соответствующей штриховкой Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
62. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком. Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.19, А— В). До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть (рис. 14.19, Г). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений. |
|
 Оба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу.
Оба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу.