|
|
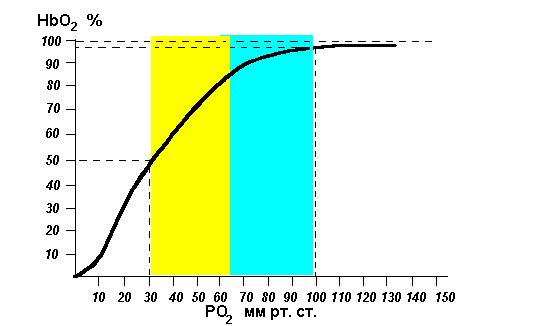
Обмен газов между кровью и тканями. Напряжение О2 и СО2 в крови, тканевой жидкости и клетках.Транспорт газов кровью. Напряжение кислорода в артериальной крови 95 мм.рт.ст. В растворенном состоянии кровью переносится всего 0,3 об.% кислорода. Основная его часть транспортируется в виде НЬО. Максимальное количество кислорода, которое может связать гемоглобин при его полном насыщении, называется кислородной емкостью крови. В норме она составляет 18-24 об.% Образование оксигемоглобина в легких и его распад в капиллярах тканей в основном обусловлены изменениями напряжения кислорода. В капиллярах легких, где напряжение его велико, происходит его образование. В тканях напряжение кислорода падает, поэтому там оксигемоглобин диссоциирует на восстановленный гемоглобин и кислород. В норме связывание гемоглобина с кислородом определяется его парциальным давлением в альвеолярном воздухе, а следовательно напряжением в крови легочных капилляров, Зависимость концентрации оксигемоглобина от напряжения кислорода в крови называется кривой диссоциации оксигемоглобина. Она не является прямо пропорциональной. При низком напряжении кислорода рост концентрации оксигемоглобина замедлен. При напряжении от 10 до 40 мм.рт.ст. он практически прямо пропорционален. А выше снова замедляется. Поэтому кривая имеет S-образную форму.
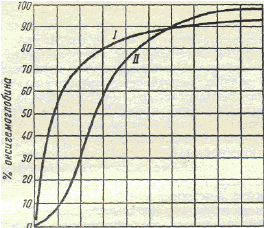
Кривые диссоциации оксигемоглобина в водном растворе (I) и в крови (II) при напряжении углекислого газа 5,33 кПа (40 мм рт. ст.) (по Баркрофту). При нулевом напряжении кислорода оксигемоглобина в крови нет. При низких значениях парциального давления кислорода скорость образования оксигемоглобина невелика. Максимальное количество гемоглобина (45— 80%) связывается с кислородом при его напряжении 3,47—6,13 кПа (26—46 мм рт. ст.). Дальнейшее повышение напряжения кислорода приводит к снижению скорости образования оксигемоглобина (рис. 25). Кроме напряжения кислорода, на образование и распад оксигемоглобина влияют и другие факторы. При сдвиге реакции крови в кислую сторону, его диссоциация ускоряется. Ее также ускоряет повышение напряжения углекислого газа и температуры. Эти изменения крови имеют место в капиллярах тканей, поэтому там они способствуют ускоренной диссоциации оксигемоглобина и освобождению кислорода. Напряжение углекислого газа в венозной крови 46 мм. рт. ст. Его перенос от тканей к легким также происходит несколькими путями. Всего в крови находится около 50 об% углекислого газа. В плазме растворяется 2,5 об.%. В виде карбгемоглобина, в соединении с глобином, переносится около 5 об%. Остальное количество транспортируется в виде гидрокарбонатов, находящихся в плазме и эритроцитах. В капиллярах тканей углекислый газ поступает в эритроциты. Там под влиянием фермента карбоангидразыон соединяется с катионами водорода и превращается в угольную кислоту. Она диссоциирует и большая часть гидрокарбонат-анионов выходит в плазму. Там они образуют с катионами натрия гидрокарбонат натрия. Меньшая их часть соединяется в эритроцитах с катионами калия, образуя гидрокарбонат калия. В капиллярах легких напряжение углекислого газа падает, а напряжение кислорода возрастает. Образующийся в эритроцитах оксигемоглобин является более сильной кислотой, чем угольная. Поэтому он вытесняет из гидрокарбоната калия анионы угольной кислоты и образует с калием калиевую соль оксигемоглобина. Освобождающиеся анионы угольной кислоты соединяются с катионами водорода. Синтезируется свободная угольная кислота. При низком напряжении углекислого газа карбоангидраза действует противоположным образом, т.е. расщепляет ее на углекислый газ и воду, которые выдыхаются. Одновременно из плазмы в эритроциты поступают анионы угольной кислоты, образующиеся в ходе диссоциации гидрокарбоната натрия. Они также образуют с катионами водорода угольную кислоту, которая расщепляется карбоангидразой на углекислый газ и воду. При дыхании из организма выводится около 200 мл углекислого газа в минуту. Это важный механизм поддержания кислотно-щелочного равновесия крови. Обмен дыхательных газов в тканях.Обмен газов в капиллярах тканей происходит путем диффузии. Этот процесс осуществляется за счет разности их напряжения в крови, тканевой жидкости и цитоплазме клеток. Как и в легких для газообмена большое значение имеет величина обменной площади, т.е. количество функционирующих капилляров. В артериальной крови напряжение кислорода 96 мм.рт.ст., в тканевой жидкости около 20 мм.рт.ст., а работающих мышечных клетках близко к 0. Поэтому кислород диффундирует из капилляров в межклеточное пространство, а затем клетки. Для нормального протекания окислительно-восстановительных процессов в митохондриях необходимо, чтобы напряжение кислорода в клетках было не менее 1 мм.рт.ст. Эта величина называетсякритическим напряжением кислородав митохондриях. Ниже ее развивается кислородное голодание тканей. В скелетных мышцах кислород накапливает белок миоглобин, по строению близкий к гемоглобину. Напряжение углекислого газа в артериальной крови 40 мм.рт.ст., в межклеточной жидкости 46 мм.рт.ст., в цитоплазме 60 мм.рт.ст. Поэтому он выходит в кровь. Количество кислорода, которое используется тканями называется коэффициентом его утилизации. В состоянии покоя ткани используют около 40% кислорода или 8-10 об%. 10(а). Транспорт газов кровью. Транспорт О2 кровью; кривая диссоциации оксигемоглобина, её характеристика; кислородная ёмкость крови; Кровь ежедневно переносит из лёгких в ткани около 600 л О2. Основной объём О2 транспортирует HbO2 (O2 обратимо ассоциирован с Fe2+ гема, это так называемый химически связанный O2 [неверный по существу, но — к сожалению — устоявшийся термин]). Незначительная часть O2 растворена в крови (физически растворённый O2). Содержание O2 в крови в зависимости от парциального давления O2 (pO2). Согласно закону Генри, количество растворённого в крови O2 пропорционально pO2 (парциальному давлению O2) и коэффициенту растворимости O2. Физическая растворимость O2 в крови примерно в 20 раз меньше, чем растворимость СО2, но для обоих газов незначительна. В то же время физически растворённый в крови газ ― необходимый этап транспорта любого газа (например, при перемещении O2 в эритроцит из полости альвеол). Кислородная ёмкость крови ― максимальное возможное количество связанного с Hb О2 ― теоретически составляет 0,062 ммоль О2 (1,39 мл О2) на 1 г Hb (реальное значение несколько меньше ― 1,34 мл О2 на 1 г Hb). Измеренные же значения составляют для мужчин 9,4 ммоль/л (210 мл О2 /л), для женщин ― 8,7 ммоль/л (195 мл О2 /л). Насыщение (сатурация, S) Hb О2 (So2) зависит от парциального давления кислорода (pO2) и фактически отражает содержание оксигенированного Hb (HbО2). So2 может принимать значения от 0 (HbО2 нет) до 1 (нет HbH). При половинном насыщении (S0,5) pO2 равно 3,6 кПа (27 мм рт.ст.), при S0,75 ― 5,4 кПа, при S0,98 13,3 кПа. Другими словами зависимость между So2 и pO2 не является линейной (характерная S-образная кривая), что благоприятствует как связыванию О2 в лёгких (артериальная кровь) и транспорту О2, так и освобождению О2 в кровеносных капиллярах органов и тканей, так как насыщение артериальной крови кислородом (SAo2) составляет примерно 97,5 %, а венозной крови (SVo2) ― 75 %. Характер насыщения таков, что кривая существенно уплощается при pO2 около 70 мм рт. ст. Так, при pO2 ниже 60 рт.ст. кислород хорошо связывается с Hb, но уже при pO2 60 мм рт. ст. насыщение составляет 90 %, и дальнейшее увеличение pO2 относительно слабо сказывается на насыщении (увеличение pO2 от 60 до 100 мм рт. ст. увеличивает насыщение всего на 7 %). Другими словами, в этом диапазоне pO2 насыщение O2 благоприятно для обеспечения его транспорта. Совершенно иная картина складывается при значениях pO2 ниже 60 мм рт. ст., то есть при небольших изменениях pO2 из Hb освобождается существенные количества O2, что облегчает его диффузию из крови в ткани. Кривая диссоциации оксигемоглобина имеет сигмоидную форму (S-образную). Это указывает на то, что субъединицы Hb работают кооперативно: чем больше О2 связывают (отдают) субъединицы, тем легче идёт ассоциация (диссоциация) последующих молекул О2. Из графика видно, что Hb (в отличие от миоглобина) имеет значительно меньшее сродство к О2 – полунасыщение гемоглобина О2 наступает при более высоком давлении О2 (около 26 мм рт. ст.).
В капиллярах покоящихся мышц, где давление О2 составляет около 40 мм рт. ст., большая часть кислорода возвращается в составе оксигемоглобина обратно в лёгкие. При физической работе pO2 в мышечных капиллярах падает до 10–20 мм рт. ст. Именно в этой области (от 10 до 40 мм рт. ст.) располагается «крутая часть» S-образной кривой, где в наибольшей степени проявляется свойство кооперативной работы субъединиц.
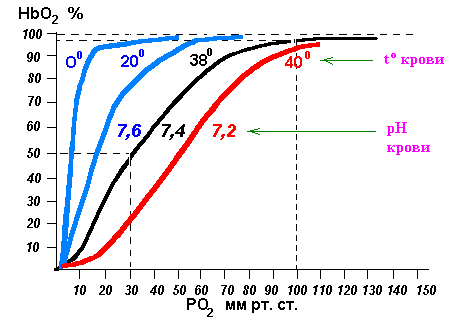
Сдвиг влево - легче насыщение кислородом:< t <Pco2> pH Сдвиг вправо - легче отдача кислорода: > t > Pco2 < pH 10.(б) Транспорт углекислоты кровью, значение карбоангидразы, взаимосвязь транспорта О2 и СО2. Углекислый газ транспортируется следующими путями: · Растворенный в плазме крови - около 25 мл / л. · Связанный с гемоглобином (карбгемоглобин) - 45 мл / л. · В виде солей угольной кислоты - букарбонаты калия и натрия в плазме крови - 510 мл / л. Таким образом, в состоянии покоя кровь транспортирует 580 мл углекислого газа в 1л. Итак, основной формой транспорта СО2 является бикорбонаты плазмы, образующихся благодаря активному протеканию карбоангидразнои реакции. В эритроцитах содержится фермент карбоангидраза (КГ), который катализирует взаимодействие углекислого газа с водой с образованием угольной кислоты, распадается с образованием бикарбонатного иона и протона. Бикарбонат внутри эритроцита взаимодействует с ионами калия, выделяемых из калиевой соли гемоглобина при восстановлении последнего. Так внутри эритроцита образуется бикарбонат калия. Но бикарбонатные ионы образуются в значительной концентрации и поэтому по градиенту концентрации (в обмен на ионы хлора) поступают в плазму крови. Так в плазме образуется бикарбонат натрия. Протон, образовавшегося при диссоциации угольной кислоты, реагирует с гемоглобином с образованием слабой кислоты ННb. В капиллярах легких эти процессы идут в обратном направлении. С ионов водорода и бикарбонатных ионов образуется угольная кислота, которая быстро распадается на углекислый газ и воду. Углекислый газ удаляется наружу. Итак, роль эритроцитов в транспорте углекислоты такова: · образование солей угольной кислоты; · образование карбгемоглобина. Диффузия газов в тканях подчиняется общим законам (объем диффузии прямо пропорционален площади диффузии, градиента напряжения газов в крови и тканях). Площадь диффузии увеличивается, а толщина диффузного слоя уменьшается при увеличении количества функционирующих капилляров, что имеет место при повышении уровня функциональной активности тканей. В этих же условиях возрастает градиент напряжения газов за счет снижения в активно работающих органах Ро2 и повышения Рсо2 (газовый состав артериальной крови, как и альвеолярного воздуха остается неизменным!).Все эти изменения в активно работающих тканях способствуют увеличению объема диффузии О2 и СО2 в них. Потребление О2 (СО2) по спирограмму определяют по изменению (сдвигу) кривой вверх за единицу времени (1 минуту). Тема 2. Регуляция дыхания. |
|