|
|
МЕДИАТОРЫ ПЕРИФЕРИЧЕСКИХ НЕРВОВВ периферической нервной системе функционируют адренергические (медиатор - норадреналин), холинергические (медиатор - ацетилхолин), пуринергические (медиаторы - АТФ и аденозин), дофаминергические и серотонинергические синапсы. Большая часть синаптотропных средств влияет на адренергические и холинергические синапсы. Различают два типа центробежных (эфферентных) нервов. • Двигательные (соматические) нервы - состоят из холинергических аксонов, следующих непрерывно от мотонейронов передних рогов спинного мозга (в составе спинномозговых нервов) или ствола головного мозга (в составе черепно-мозговых нервов) к скелетным мышцам. • Вегетативные нервы - иннервируют внутренние органы. Различают симпатические и парасимпатические вегетативные нервы. Симпатические нервные пути состоят из коротких преганглионарных и длинных постганглионарных волокон, образующих синаптический контакт в паравертебральных вегетативных ганглиях (симпатический ствол). Центры преганглионарных волокон находятся в боковых рогах грудного и поясничного отделов спинного мозга (сегменты с C8 по L3). Симпатические преганглионарные волокна - холинергические; постганглионарные - адренергические. Парасимпатические нервные пути состоят из длинных преганглионарных и коротких постганглионарных волокон. Ганглии локализованы около исполнительных органов или интрамурально. Парасимпатические волокна входят в состав некоторых черепно-мозговых нервов (глазодвигательного, лицевого, языкоглоточного, блуждающего) и спинномозговых тазовых нервов. Парасимпатические центры спинного мозга находятся в боковых рогах крестцовых сегментов S2-S4. Парасимпатические преганглионарные и постганглионарные волокна - холинергические. Эфферентные волокна, иннервирующие мозговой слой надпочечников, не прерываются в вегетативных ганглиях и, по сути, являются преганглионарными холинергическими волокнами. Таким образом, хромаффинную ткань надпочечников рассматривают как аналог симпатических ганглиев, у которых редуцировались аксоны, а медиатор стал гормоном (70-90% составляет адреналин, 10-30% - норадреналин). В синокаротидной зоне холинорецепторы расположены на окончаниях каротидной ветви языкоглоточного нерва, тонизирующей дыхательный центр, ацетилхолин выделяется из синаптических пузырьков клеток каротидных клубочков. Таким образом, адренергические волокна - только симпатические постганглионарные; холинергические волокна - двигательные, симпатические преганглионарные, парасимпатические преганглионарные и постганглионарные. АДРЕНЕРГИЧЕСКИЕ СИНАПСЫ Адренергические нейроны расположены в ЦНС (голубое пятно среднего мозга, мост, продолговатый мозг) и симпатических ганглиях. Периферические адренергические синапсы образованы варикозными утолщениями разветвлений постганглионарных симпатических волокон. Медиатором адренергических синапсов является норадреналин. Его биохимический предшественник дофамин выполняет медиаторную функцию в дофаминергических синапсах. Адреналин - гормон мозгового слоя надпочечников. Все три вещества относятся к группе катехоламинов, так как содержат гидроксильные группы в 3-м и 4-м положении ароматического кольца. Синаптические пузырьки в адренергических синапсах по данным электронной микроскопии имеют гранулярное строение и поэтому получили название «гранулы». В гранулах норадреналин депонирован в связи с АТФ и белком хромогранином. В составе гранул обнаружены также ферменты и модулирующие нейропептиды (энкефалины, нейропептид Y). Норадреналин синтезируется из аминокислоты тирозина. Превращение фенилаланина в тирозин является неспецифическим процессом и происходит в печени. Обе аминокислоты содержатся в больших количествах в твороге, сыре, шоколаде, бобовых. Тирозин активным транспортом поступает в адренергические окончания. В их аксоплазме он приобретает второй гидроксил-радикал в 3-м положении ароматического кольца и превращается в диоксифенилаланин (ДОФА). Эту реакцию катализирует тирозингидроксилаза митохондрий. Затем ДОФА декарбоксилазой ароматических L-аминокислот декарбоксилируется в дофамин. Дофамин из аксоплазмы транспортируется в гранулы. На последнем этапе дофамин приобретает третий гидроксил в β-положении боковой цепи при участии дофамин- β-гидроксилазы. В мозговом слое надпочечников норадреналин выходит из гранул и в цитоплазме метилируется в гормон адреналин под действием N-метилтрансферазы (донатором метильных групп служит S-аденозил-метионин) (рис. 9-1). Образование адреналина повышают глюкокортикоиды, эстрогены и тироксин. Глюкокортикоиды, поступая в мозговой слой по воротной системе надпочечников, активируют тирозингидроксилазу, дофамин- β-гидроксилазу и N-метилтрансферазу. У некоторых видов акул корковый и мозговой слои надпочечников представляют собой изолированные железы, поэтому у них адреналин не синтезируется, а единственным гормоном хромаффинных клеток является норадреналин. После диссоциации комплекса «норадреналин-адренорецептор» медиатор инактивируется в течение 2-3 мин при участии ряда механизмов. • Нейронального захвата (захвата-1) - активного транспорта вначале через пресинаптическую мембрану (сопряжен с выходом ионов натрия), а затем через мембрану гранул под влиянием протонной АтФазы (при входе в гранулу одной молекулы норадреналина в аксоплазму выходят 2 протона). • Экстранейронального захвата (захвата-2) нейроглией, фибробластами, кардиомиоцитами, клетками эндотелия и миоцитами сосудистой стенки. • Инактивации ферментами.
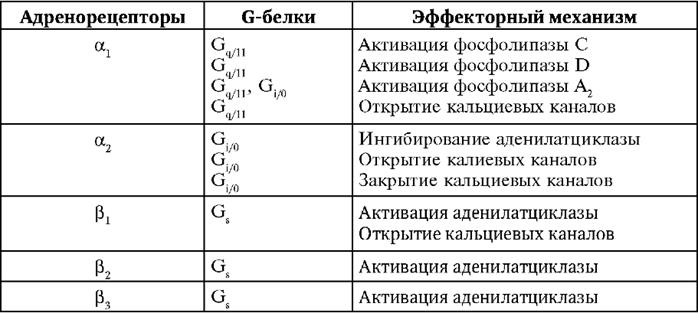
Рис. 9-1.Биосинтез адреналина Около 80% норадреналина подвергается нейрональному захвату и по 10% - экстранейрональному захвату и ферментативному расщеплению. Необходимость нейронального захвата диктуется дефицитом субстратов и большой потребностью в энергии для синтеза норадреналина из тирозина. Для сохранения адреналина основное значение имеет экстранейрональный захват. Ферменты инактивации катехоламинов - МАО и катехол-О-метил-трансфераза (КОМТ). МАО, локализованная на внешней мембране митохондрий и в гранулах, осуществляет окислительное дезаминирование катехоламинов с образованием биогенных альдегидов. Затем альдегиды окисляются НАД-зависимой альдегиддегидрогеназой в кислоты или восстанавливаются альдегидредуктазой в гликоли. Цитоплазматический фермент КОМТ катализирует присоединение метильной группы к гидроксилу в 3-м положении ароматического кольца (только при наличии гидроксила в 4-м положении). Донатором метильных групп служит S-аденозилметионин. Метилированные продукты в 200-2000 раз (по результатам разных тестов) менее активны, чем норадреналин и адреналин. АДРЕНОРЕЦЕПТОРЫ В 1948 г. английский фармаколог Рассел Алквист выдвинул гипотезу о двух типах адренорецепторов. α-Адренорецепторы суживают сосуды, наиболее чувствительны к эпинефрину, меньше реагируют на норэпинефрин и очень слабо воспринимают действие изопреналина (изопропилнорадреналина). β-Адренорецепторы расширяют сосуды, обладают максимальной чувствительностью к изопреналину, в 10-50 раз слабее реагируют на эпинефрин и норэпинефрин. Адренорецепторы обнаружены на постсинаптической, пресинаптической мембранах и в клетках, не получающих адренергической иннервации. Постсинаптические адренорецепторы имеют индексы 1 или 2, пресинаптические и внесинаптические адренорецепторы обозначаются индексом 2. Внесинаптические адренорецепторы активируются циркулирующими в крови норадреналином и адреналином. β-Адренорецепторы жировой ткани имеют индекс 3. Все адренорецепторы характеризуются сходной последовательностью аминокислот (у α1- и α2-адренорецепторов идентичны 30% аминокислот, у β1- и β2-адренорецепторов - 60%). В сосудах и внутренних органах расположены α- и β-адренорецепторы различных типов. Например, в сосудах легких обнаружено 30% β1-адренорецепторов и 70% β,,-адренорецепторов. Адренорецепторы являются гликопротеинами, ассоциированы с G-белками и имеют такое же строение, как и другие рецепторы этого типа. Их белковая цепь состоит из семи гидрофобных доменов в виде трансмембранной спирали, Домены соединены гидрофильными петлями, расположенными попеременно по обе стороны мембраны. N-конец белковой молекулы адренорецептора расположен внеклеточно, C-конец - внутри клетки. Активный центр адренорецептора представляет собой карман, образованный высококонсервативными аминокислотами, расположенными в средней и во внеклеточной третях гидрофобных трансмембранных спиралей. Аминогруппа катехоламинов соединяется ионной связью с карбоксилом аспарагиновой кислоты в третьем трансмембранном домене. Гидроксилы катехоламинов образуют водородную связь с остатками серина в пятом и седьмом доменах, что необходимо для активации адренорецепторов. Сведения о механизмах функционирования, чувствительности к агонистам и антагонистам, физиологической роли адренорецепторов представлены в табл. 9-2-9-4. Таблица 9-2.Адренорецепторы и их эффекторные системы
α-Адренорецепторы Постсинаптические α1-адренорецепторы (типы A, B, D) активируют мембранные фосфолипазы и увеличивают проницаемость кальциевых каналов. В гладких мышцах ионы кальция активируют Таблица 9-3.Адренорецепторы
Примечания. Э - эпинефрин; НЭ - норэпинефрин; И - изопреналин. Таблица 9-4.Влияние вегетативной нервной системы на функции эффекторных органов
Продолжение табл. 9-4
Продолжение табл. 9-4
Окончание табл. 9-4
* В гладких мышцах и железах находятся различные типы м-холинорецепторов (преобладают м-холинорецепторы 3-го типа), в сердце локализованы м2-холинорецепторы. ** Указана сила влияния на соответствующие функции. *** В сердце преобладают β1-адренорецепторы. **** Указаны преобладающие типы адренорецепторов, в артериолах органов брюшной полости и почек находятся сосудорасширяющие рецепторы дофамина. кальмодулинзависимую киназу легких цепей миозина, что необходимо для образования актомиозина и сокращения. Только в желудке и кишечнике α1-адренорецепторы, открывая кальцийзависимые калиевые каналы, вызывают гиперполяризацию сарколеммы и расслабление гладких мышц. Эффекты активации α1-адренорецепторов следующие: • сокращение радиальной мышцы радужки с расширением зрачков (мидриаз, от греч. amydros - «темный, неясный»); • сужение сосудов кожи, слизистых оболочек, органов пищеварения, почек и головного мозга; • повышение АД; • сокращение капсулы селезенки с выбросом депонированной крови в циркуляторное русло; • сокращение сфинктеров пищеварительного тракта и мочевого пузыря; • подавление моторики и снижение тонуса желудка и кишечника. α2-Адренорецепторы (типы A, B, C) локализованы на постсинаптической, пресинаптической мембранах и вне синапсов. Пресинаптические α2-адренорецепторы ингибируют аденилатциклазу и тормозят синтез цАМФ, увеличивают проницаемость мембран для K+ с развитием гиперполяризации, блокируют кальциевые каналы. По принципу отрицательной обратной связи они тормозят выделение норадреналина из адренергических окончаний при избыточной активации адренорецепторов. Постсинаптические α2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, подавляют секрецию кишечного сока. Внесинаптические α2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, секрецию инсулина, повышают агрегацию тромбоцитов. β-Адренорецепторы β-Адренорецепторы, активируя аденилатциклазу, вызывают превращение АТФ в цАМФ - активатор цАМФ-зависимых протеинкиназ. Протеинкиназа А транспортируется в ядро клеток и фосфорилирует фактор транскрипции - ДНК-связывающий белок. Этот белок регулирует активность цАМФ-чувствительного элемента в промоторном участке гена, в результате активируется синтез β-адренорецепторов. Для постсинаптических β1-адренорецепторов характерны следующие эффекты: • стимуляция сердечной деятельности: тахикардия, ускорение проведения возбуждения по проводящей системе, усиление сокращений миокарда, увеличение потребности в кислороде (β1-адренорецепторы при участии Gs-белков стимулируют фосфорилирование кальциевых каналов, что сопровождается их открытием, входом в саркоплазму Ca2+ и мобилизацией этого иона из саркоплазматического ретикулума; также фосфорилируется белок фосфоламбан, способствующий депонированию Ca2+ в саркоплазматическом ретикулуме); • стимуляция секреции ренина; • угнетение моторики кишечника. Постсинаптические и внесинаптические β2-адренорецепторы расслабляют гладкие мышцы и вызывают гипергликемию. В гладких мышцах они при участии цАМФ снижают активность киназы легких цепей миозина, в печени и скелетных мышцах активируют фермент гликогенолиза фосфорилазу, ингибируют гликогенсинтазу. Кроме того, в гладких мышцах блокируются кальциевые каналы и активируются калиевые каналы. Типичные эффекты β2-адренорецепторов следующие: • расширение сосудов сердца, легких и скелетных мышц; • снижение АД; • расширение бронхов и угнетение секреторной функции бронхиальных желез; • угнетение моторики желудка и кишечника; • расслабление желчного пузыря, мочевого пузыря, матки; • усиление цАМФ-зависимого гликогенолиза и глюконеогенеза; • активация гликогенолиза в скелетных мышцах; • стимуляция секреции инсулина. Пресинаптические β2-адренорецепторы осуществляют обратную связь, стимулируя выделение норадреналина при недостаточной активации адренорецепторов. β3-Адренорецепторы усиливают цАМФ-зависимый липолиз в жировых депо с повышением в крови содержания свободных жирных кислот. Полиморфизм гена β3-адренорецептора создает предрасположенность к развитию ожирения и сахарного диабета 2-го типа. β-Адренорецепторы подвергаются гомологической и гетерологической десенситизации. Гомологическая десенситизация развивается только к действию β-адреномиметиков. Под их влиянием медленно фосфорилируется специфическая цАМФ-независимая протеинкиназа - киназа β-адренорецепторов. Этот фермент фосфорилирует комплекс "β-адренорецепто-β^-адреномиметик". В дальнейшем присоединяется белок β-аррестин, нарушающий связь регуляторного домена β-адренорецепторов с G-белком. Инактивированный комплекс подвергается интернализации в цитоплазму и деградации в лизосомах. При гетерологической десенситизации в ответ на действие β-адреномиметиков ослабляется реакция на многие вещества, повышающие синтез цАМФ. Она обусловлена быстрым (в течение миллисекунд) фосфорилированием β-адренорецептора протеинкиназой А с последующей утратой способности передавать сигнал на эффекторные системы. |
|