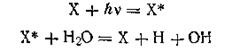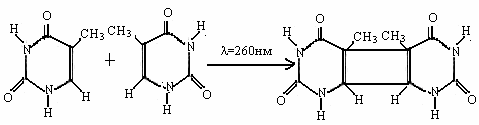|
|
Фотохимические процессы в биологических системах
Из всех известных в природе фотохимических процессов наибольшее значение имеет фотосинтез. Основоположником учения о фотосинтезе является К. А. Тимирязев. Фотосинтез является основой существования всего живого на земле. Фотосинтез зеленых растений — это единственный первоисточник накопления органического вещества на Земле, которое служит для питания человека и животных. Вся растительность земного шара создает ежегодно около 120 млрд. т органического вещества, из них примерно 10 млрд. тонн производит человек, выращивая на площади около 2,5 млрд. га пищевые и кормовые растения. Кислород, содержащийся в атмосфере Земли, имеет биогенное происхождение, т. е. также является продуктом фотосинтеза зеленых растений. Расчет показывает, что растениями ежегодно возвращается в атмосферу около 4,7.1011 т газообразного кислорода. В зеленом листе растения под воздействием солнечной радиации протекает целый комплекс фотохимических процессов, в результате которых из воды, углекислого газа и минеральных солей образуются крахмал, клетчатка, белки, жиры и другие сложные органические вещества. Процесс фотосинтеза очень сложен. Он осуществляется при непосредственном участии важнейшего природного фотокатализатора—хлорофилла и сопровождается целым циклом химических превращений, не зависящих от солнечной радиации. В этих превращениях участвует большое число разнообразных биокатализаторов — ферментов. Суммарное уравнение фотосинтеза обычно выражают в виде реакции превращения двуокиси углерода и воды в гексозу: 6СО2 + 6Н2О = С6Н12О6 + 6О2 Однако это уравнение, как и большинство суммарных уравнений в биологии, не выражает основных особенностей процесса. Важнейшей заслугой К. А. Тимирязева является материалистическое научное обоснование фотосинтеза. Тимирязев впервые показал, что фотосинтез подчиняется закону сохранения и превращения энергии. Таким образом, были опровергнуты идеалистические воззрения на процесс фотосинтеза, объяснявшие его действием нематериальной «жизненной силы». Не менее важной заслугой Тимирязева является открытие роли хлорофилла как сенсибилизатора фотохимических реакций, происходящих при фотосинтезе. Он экспериментально установил, что фотосинтез осуществляется преимущественно в красных и синих лучах видимого спектра. Тимирязев провел следующий опыт. Ряд стеклянных трубочек, наполненных смесью воздуха и диоксида углерода и содержащих по одному одинаковому зеленому листу, был выставлен на разложенный с помощью трехгранной призмы солнечный свет так, что в каждой части солнечного спектра находилась одна трубочка. Через каждые несколько часов определялось содержание диоксида углерода в трубочках. Оказалось, усвоение СО2 происходит только в тех лучах, которые поглощаются хлорофиллом, т. е. в красных, оранжевых и желтых частях спектра. Таким образом, Тимирязев показал, что именно хлорофилл является поглотителем света в зеленых растениях и что этот пигмент, поглощая кванты света, обладает способностью передавать их далее молекулам веществ, являющихся исходными при фотосинтезе.
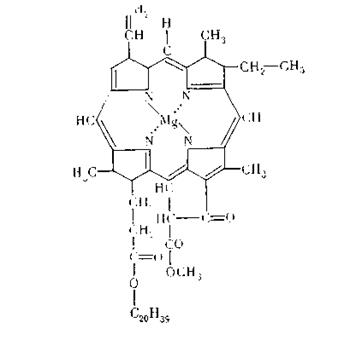
При этих реакциях хлорофилл испытывает обратимое окислительно-восстановительное превращение. В основеструктуры хлорофилла лежит порфириновое ядро, называемое хлорином. Оно состоит из четырех соединенных СН-мостиками остатков пиррола, которые связаны двумя основными и двумя координационными связями с центральным атомом магния. Кроме того, в молекулу хлорофилла входит остаток молекулы высокомолекулярного непредельного спирта фитола. В настоящее время известно не менее пяти видов хлорофилла, которые отличаются друг от друга строением молекулы. Помимо хлорофилла, который является основным видом фотосинтетических пигментов, в зеленом листе (в так называемых хлоропластах, представляющих собой сложные специализированные биологические структуры) содержатся и другие пигменты — каротиноиды и фикобелины, которые обычно называют вспомогательными. Эти пигменты, по современным представлениям, принимают известное участие в фотосинтезе, а также защищают хлорофилл от фотоокисления. Помимо пигментов, основными компонентами хлоропластов, в которых, собственно, и осуществляется весь процесс фотосинтеза, являются липоидные вещества и белки, которые содержат большое число ферментов, необходимых для осуществления последующих стадий фотосинтеза, не связанных с воздействием солнечной радиации. Многие вопросы фотосинтеза, несмотря на бурное развитие науки, остаются мало изученными и до настоящего времени. Как уже упоминалось ранее, процесс фотосинтеза состоит из двух стадий — световой и темновой, причем обе эти стадии тесно связаны между собой. Поскольку исходным процессом фотосинтеза является поглощение света хлорофиллом, приближенно фотосинтез можно представить в виде следующей схемы. В световой стадии хлорофилл, поглотив квант света, переходит в возбужденное состояние и в таком виде через ряд промежуточных процессов вызывает разложение молекулы воды на атом водорода Н и радикал ОН по схеме
где символом X условно обозначена молекула хлорофилла; X* — та же молекула в активном состоянии. Далее молекула хлорофилла, присоединяя атом водорода, восстанавливается. Радикалы ОН, соединяясь попарно, образуют молекулу пероксида водорода Н2О2, которая как непрочное соединение распадается на воду и кислород: X + Н = ХН 4ОН = 2Н2О2 2Н2О2 = 2Н2О + О2 После завершения этих реакций наступает темновая стадия процесса фотосинтеза, сущность которой состоит в передаче водорода молекулой восстановленного хлорофилла молекуле СО2 с образованием органических соединений типа углеводов. Этот процесс совершается под действием соответствующих ферментов по схеме: 4Н + СО2 = СН2О + Н2О В итоге за счет полимеризации получается конечный продукт фотосинтеза — гексоза С6Н12О6. То, что выделяемый в процессе фотосинтеза кислород принадлежит воде, а не двуокиси углерода, было доказано А. П. Виноградовым (1946) с помощью метода меченых атомов. Так, при использовании воды Н218О весь ее кислород 18О был найден после осуществления фотосинтеза в свободном молекулярном кислороде, а при работе с С18О2 и Н2160 выделяется свободный кислород 16О, тогда как кислород 18О входил в состав органических соединений. Установление этого факта имело существенное значение для теории фотосинтеза, так как ранее многие ученые считали, что молекулярный кислород образуется путем светового разложения или фотолиза СО2. Приведенная выше схема фотосинтеза является лишь приближенной и не отражает всех деталей этого чрезвычайно сложного явления. В последние годы было установлено, что на восстановление одной молекулы СО2 до углерода затрачивается не один, а 8—12 квантов энергии. Это свидетельствует о том, что в процессе фотосинтеза происходит по крайней мере восемь первичных фотохимических реакций, которые совершаются в определенном порядке с другими (не фотохимическими) реакциями. Известно, что далеко не каждая молекула хлорофилла или другого пигмента, поглотившая свет и сохранившая достаточное количество энергии для фотохимической реакции, является центром подобной реакции. На самом деле фотохимическая активность, т. е. непосредственная связь с фотохимической реакцией, осуществляется лишь примерно одной молекулой из 200—250 молекул хлорофилла. Об этом явлении А.Г. Пасынский пишет: «...Могло бы создаться неправильное представление, что основная масса хлорофилла является фотохимически неактивной и играет в листе роль запасного вещества, как иногда предполагалось в литературе. В действительности такое положение является необходимым следствием квантовой природы действующего света. Поглощение света данной молекулой хлорофилла не происходит непрерывным потоком; кванты света, падающие подобно каплям дождя, поглощаются все время разными молекулами хлорофилла. По данным Рабиновича, даже на прямом солнечном свету каждая молекула хлорофилла поглощает квант света всего один раз за 0,1 с, а при менее благоприятных условиях — гораздо реже. Между тем скорость последующих ферментативных реакций является чрезвычайно высокой. Если бы в этих условиях каждая молекула хлорофилла была самостоятельным центром фотохимической реакции, связанным с необходимыми вспомогательными ферментами, то такое устройство было бы столь же нецелесообразно, как если бы каждый участок крыши, на который падает отдельная капля дождя, был оборудован самостоятельным водостоком. В листе для подобного устройства просто не хватило бы места, не говоря уже о том, что оно могло бы использоваться лишь незначительную часть времени. Напротив, соединение большой группы (200—250) молекул хлорофилла с одним центром фотохимической реакции обеспечивает его непрерывную работу, подобно тому, как присоединение одного водостока к достаточно значительной поверхности крыши позволяет получить из отдельных капель непрерывный поток воды. Ясно, что при этом вся масса молекул хлорофилла активно участвует в полезномпроцессе, хотя она связана лишь с одним центром превращения поглощенной лучистой энергии в химическую». Все это лишний раз подтверждает чрезвычайную сложность процесса фотосинтеза, каждая ступень которого требует не только определенных условий среды, но и очень сложной системы вспомогательных веществ, а также строго определенной внутренней структуры внутриклеточного содержимого. На важность структурных факторов указывает то, что зеленый лист, подвергшийся механическому воздействию (например, если прокатать его на стекле толстой стеклянной палочкой), теряет способность к фотосинтезу. Изучение процессов фотосинтеза очень важно не только с чисто теоретической точки зрения, но и с точки зрения получения высоких и устойчивых урожаев. Познать эти процессы, научиться управлять ими — вот те задачи, на решение которых направлены в настоящее время усилия целой армии отечественных и зарубежных ученых. Очень важное значение имеют происходящие под действием УФ - света реакции димеризации азотистых оснований в ДНК и РНК. Основными хромофорами (хромофор - часть молекулы, поглощающая свет и обусловливающая цвет вещества) молекул ДНК являются азотистые основания нуклеотидов. Поглощение азотистыми основаниями квантов УФ – света приводит к образованию электронно-возбужденных синглетных состояний, возникающих в результате П → П* переходов.
В электронновозбужденном состоянии пиридиновые основания вступают в реакцию димеризации, которая состоит в соединении двух азотистых оснований по 5,6 - двойной углеродной связи с образованием циклобутанового кольца между остатками молекул азотистых оснований. Таким образом, отдельные нуклеотиды оказываются связанными не только через остатки фосфорной кислоты, но и через азотистые основания. Для этой реакции квантовый выход γ = 2 ∙10-2 . Эта реакция вызывает так называемые "точечные" мутации; 80 % всех летальных мутаций, связанных с результатами воздействия УФ - излучения, являются следствиями димеризации тимина.
При небольшой интенсивности облучения возникают полезные точечные мутации. В результате облучения родительских форм УФ - светом и отбора полезных признаков создан сорт пшеницы Эритроспермум-103.
|
|