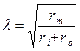|
|
Кабельные свойства нервных проводниковЕсли нерв или мышцу длительно подвергать действию постоянного тока, можно обнаружить характерные закономерности изменения ее возбудимости во времени. Приложение тока к возбудимой ткани требует использования двух электродов, катода и анода. Если ток течет через ткань от анода к катоду, под катодом уровень мембранного потенциала уменьшается (деполяризация) и приближается к КУД. Под анодом происходит обратный процесс, гиперполяризация. При замыкании цепи возбуждение возникает под катодом, при размыкании – под анодом. Это и составляет существо «полярного закона». В современной интерпретации формулировка «полярного закона» возбуждение возбудимой ткани или клетки достигается только тем током, который вызывает деполяризацию мембраны лучше отражает происходящие события. В основе рассматриваемых явлений лежит способность мембраны дополнительно накапливать электрический заряд из-за своей электрической емкости. В классической электрофизиологии при действии постоянного тока на ткань повышение возбудимости под катодом называют катэлектротоном, а понижение ее под анодом — анэлектротоном. Катэлектротон и анэлектротон вместе объединяют как электротонические явления. В них проявляется возможность управления возбудимостью тканей организма при помощи подпорогового электрического тока. При длительном воздействии тока на ткань ее возбудимость может чрезвычайно измениться так, что катэлектротон превратиться в катодическую депрессию (возбудимости под катодом снизится), а анэлектротон в анодическую экзальтацию (будет наблюдаться повышением возбудимости под анодом). В основе рассматриваемых явлений лежит изменение свойств потенциалзависимых ионных каналов под действием постоянного электрического тока. При катодической депрессии обнаружено ослабление проницаемости потенциалзависимых натриевых каналов и усиление проницаемости калиевых каналов для соответствующих ионов. Смысл закона «все или ничего» состоит в том, что при возбуждении нервных волокон или тканей, построенных по принципу функционального синцития, любое сверхпороговое раздражение вызывает сразу полный ответ (ПД или сокращение). Этот закон обусловлен независимостью потенциалзависимых натриевых и кальциевых каналов от других параметров, кроме степени поляризации мембраны (величины мембранного потенциала), КУД и порога. Закон крутизны раздражения, или закон градиента обусловлен необходимостью существования определенного времени, в течение которого может достигаться конечное значение порогового раздражения. В условиях, при которых деполяризующий ток сообщается возбудимой клетке не мгновенно, а плавно, потенциал действия может не возникнуть, хотя формально напряжение между полюсами раздражающего электрода достигнет высокого, порогового в случае прямоугольного по форме стимула, значения. Оказывается, критический уровень деполяризации при постепенном сдвиге напряжения на мембране при пологом стимуле не достигается из-за частичной инактивации натриевых каналов и активации калиевых каналов утечки. Мембранный потенциал «не догоняет» все возрастающий новый уровень КУД. Закон градиента демонстрирует одно из проявлений феномена аккомодации. Возбудимые клетки по способности к мембранной аккомодации различаются. Аккомодация чувствительных нервных волокон низка, поэтому при длительном смещении мембранного потенциала до КУД они способны генерировать множество ПД, частота которых определяется в этом случае рефрактерностью. Напротив, двигательные нервные волокна, управляющие мышцами, имеют высокую способность к аккомодации из-за медленной активации их калиевых каналов при деполяризации. Другое проявление аккомодации – закон сила-длительность, или закон гиперболы. В общем виде он может быть определен как феномен обратной зависимости между силой действующего раздражителя и ответной реакцией возбудимой ткани (но не отдельной клетки!). Графически закон описывается кривой обратной пропорциональной зависимости в соответствии с формулой Qпор=Iпор× t; Iпор= Qпор/ t В эксперименте и клинических исследованиях определяют действие закона применением разных периодов стимуляции и разных напряжений стимулирующего тока, вызывающего пороговое возбуждение объекта. Первым определяемым параметром является время, t. Начиная от заведомо избыточной, постепенно уменьшают длительность стимула, каждый раз определяя порог появления реакции. Так определяют полезное время – наименьшую длительность стимула, при которой наблюдается одинаковый порог реакции. Эту величину называют реобаза. На втором этапе определяют длительность стимула, вызывающего пороговое возбуждение объекта при фиксированном значении напряжения стимуляции в 2 реобазы. Минимальная длительность прямоугольного электрического импульса с амплитудой, равной 2 реобазам, вызывающего пороговое возбуждение, называется хронаксией. Хронаксия отражает возбудимость системы, являясь ее обратной величиной – чем больше хронаксия, тем меньше возбудимость объекта. По отдельным нервным волокнам сигналы распространяются в виде потенциалов действия и электротонических потенциалов, но на разные предельные расстояния. Способность аксонов и дендритов, а также мембран мышечных клеток проводить электрические сигналы характеризуется их кабельными свойствами. Именно эти свойства определяют и условия регистрации внеклеточных потенциалов, или электрографию. Кабельные свойства нервных проводников существенны для распространения сигналов в нервной системе. Они обусловливают генерацию потенциалов действия в сенсорных нервных окончаниях, или рецепторах, при действии раздражителей, проведение сигналов по дендритам и аксонам, суммацию сигналов мембраной перикариона нейрона. Постоянная длины λ, или пространственная константа поляризации зависит от сопротивления мембраны rm, сопротивления внешней среды ro и сопротивления аксоплазмы ri.
Чем больше сопротивление мембраны и меньше сопротивление среды, тем на более протяженное расстояние передается электротонический потенциал. Уменьшение величины электротона в зависимости от расстояния называется декрементом. Оказалось, что на постоянную длины влияет и диаметр проводника, поскольку от него зависит сопротивление аксоплазмы. Поэтому толстые нервные волокна имеют большее расстояние λ, на которое способен распространяться электротон. Распространение электротона – прежде всего катэлектротона – важный физиологический феномен. В клетках, не генерирующих ПД (глия, эпителий, тонические мышечные волокна), за счет проведения электротона осуществляется функциональная взаимосвязь между клетками. В дендритном дереве нейронов, например, коры больших полушарий мозга, сигналы в виде электротона могут доходить от дендритов до сомы. В области синапса, зная рассмотренные закономерности, можно определить, на какое расстояние могут распространяться синаптические потенциалы. Электротон способствует передаче возбуждения между кардиомиоцитами и проведению ПД. Тем не менее, и расчеты, и прямые измерения дают очень небольшие расстояния, на которое способен распространяться электротонический сигнал. Поэтому, если деполяризация в участке мембраны не достигает точки КУД, нет увеличения проницаемости для Na+ через потенциал-зависимые каналы, происходят только пассивные изменения мембранного потенциала. Константа λ варьирует от 0,1 до 5 мм. Однако в некоторых случаях пространственная константа поляризации имеет подчеркнуто важное значение. Например, при распространении фронта возбуждения в миокарде λ предопределяет синхронное прохождение возбуждения по толще кардиомиоцитов. Важна эта константа и при распространении потенциала поля по гладким мышцам в кишечнике. Рассмотрим события, связанные с распространением ПД по немиелинизированному нервному волокну (иногда с ошибкой говорят немиелиновому, но это бессмысленно, так как миелин не проводит ток, это изолирующая проводник оболочка!). Инициация волны возбуждения может быть обусловленной либо активностью рецептора (генераторный потенциал), либо синаптическими процессами. Можно вызвать ПД и электростимуляцией аксона. Если локальное смещение мембранного потенциала (деполяризация) превышает порог, достигает точки КУД, активирует потенциалзависимые натриевые каналы, ПД в виде волны деполяризации-реполяризации возникает и распространяется вдоль нервного волокна. Точка максимума ПД соответствует максимальной реверсии мембранного потенциала (овершут). Получается ситуация, при которой ПД, распространяясь по волокну, порождает перед собой токи Германна, которые разряжают мембранную емкость, приближают мембранные потенциал следующего участка мембраны к КУД, и т.д. Сзади за собой перемещающийся ПД оставляет область мембраны, находящуюся в состоянии относительной рефрактерности. Для распространения ПД необходимо, чтобы он всякий раз порождал электротонический потенциал в соседней области, той, куда он распространяется, способный сместить мембранный потенциал на величину порога, то есть амплитуда потенциала действия должна превышать порог его возникновения в несколько раз. Отношение ПД/порог носит название гарантийный фактор (Uпд/Uпорог=5..7). Скорость перемещения электротона равна скорости распространения электромагнитных волн (ведь электротон есть просто электрический ток, переносимый ионами, прежде всего ионами калия из-за их избытка в цитозоле, только с высокой степенью утечки). Скорость же перемещения ПД по безмякотным волокнам мала и не превышает 1 м/с. У кальмара, за счет объединения в эмбриогенезе нескольких аксонов в один, что увеличивает общий диаметр проводника, скорость импульса в немиелинизированном волокне может быть до 25 м/с. У млекопитающих скорость повышена за счет миелинизации аксонов. Высокое удельное сопротивление миелина приводит к тому, что мембрана мякотных волокон приобретает высокое сопротивление и малую емкость. В перехватах Ранвье сосредоточены натриевые потенциалзависимые каналы, в приперехватных областях – калиевые, ответственные за реполяризацию. Эти особенности строение приводят к тому, что сальтаторное проведение возбуждения имеет высокую надежность и высокую скорость, которые сочетаются с экономичностью (мякотные аксоны для перемещения Na+ и K+ через мембрану обходятся меньшим количеством натрий-калиевой АТФазы). Отличительное биофизическое свойство сальтаторного проведения ПД состоит и в том, что токи замыкаются через межклеточную среду, имеющую низкое сопротивление, при этом токи следуют и вдоль, и поперек волокна. Скорость передачи импульсов по мякотному волокну зависит от диаметра последнего простым соотношением V=К×d, где d –диаметр, а к-константа. Для амфибий к=2, для млекопитающих к=6. Длина участка волокна, вовлеченного в процесс передачи одного ПД равна L=t×V, где t-длительность импульса. Этот показатель важен в методическом отношении, поскольку от длины возбужденного участка нерва зависит подбор межполюсного расстояния отводящих (регистрирующих) электродов. В нервных стволах отдельные афферентные и двигательные нервные волокна расположены в компактно упакованном состоянии. Проведение по отдельным волокнам осуществляется изолированно от соседних, может распространяться в двух направлениях от места возникновения, имеет относительно постоянную скорость в любом участке аксона (кроме окончаний) и возбуждение от нескольких источников возникновения в клетке может подвергаться алгебраическому суммированию. Диапазон различий в скоростях проведения в волокнах велик, что позволило провести несколько классификаций. Наиболее принятыми считаются классификации Эрлангера-Гассера (группы АαβγδВС) и, в меньшей степени, Ллойда –Макинтайра (группы I, II, III и IV) [25].
|
|